NEOGENE PLANKTONIC FORAMINIFERAL STUDIES
Neogene planktonic foraminifers in the southern Indo-Pacific region were studied notably by Jenkins (1971, 1985; mainly from New Zealand), Brönnimann and Resig (1971; southwest Pacific, Deep Sea Drilling Project [DSDP] Leg 7), Jenkins and Srinivasan (1986; southwest Pacific, DSDP Leg 90), Srinivasan and Kennett (1981; South Pacific), Kennett et al. (1985; Indo-Pacific), Wright and Thunell (1988; Indian Ocean), Huber (1991; Kerguelen Plateau, Ocean Drilling Program [ODP] Leg 119), Berggren (1992; Kerguelen Plateau, ODP Leg 120), and Chaisson and Leckie (1993; equatorial western Pacific). In southern Australia, McGowran et al. (1971) first attempted the correlation of local assemblages to tropical zonations and McGowran (1986) emphasized species datum levels while reviewing planktonic foraminiferal biostratigraphy of the southern Indo-Pacific region. McGowran and Li (1997) reported their detailed study on the Lakes Entrance section in southeastern Australia, from which Jenkins (1960) pioneered a local planktonic foraminiferal zonation.
In southern Australia, the (sub)tropical zonations of Blow (1979) and Berggren et al. (1995) cannot be directly recognized because of the rarity or absence of many zonal markers. The southern mid-latitude zonation of Jenkins (1993) (Fig. F2) appears to be relevant, but three reasons prevented its use in this study: (1) lack of chronologic constraint on many datum levels, (2) difficulties in correlating it with other schemes (see also Hornibrook et al., 1989), and (3) at least some datum levels probably from a more incomplete stratigraphy than previously realized. Examples of the latter two points include the FO of Globoconella puncticulata, which has been used to date the Miocene/Pliocene boundary by Jenkins (1971, 1993; as Globorotalia puncticulata). Jenkins (1993) also recognized the overlying Zone SN12 (including Globorotalia pliozea and Globorotalia puncticulata Subzones) as representing the early Pliocene, with the top defined by the LO of Globorotalia inflata. The succession, as summarized in Jenkins (1993), from the FO of G. puncticulata (also FO of G. pliozea) to the LO of G. pliozea to the FO of G. inflata could not be ratified by Hornibrook et al. (1989, p. 144), who showed that G. pliozea ranged as high as the LO of Globorotalia margaritae. Based on a magnetostratigraphy, Berggren et al. (1995) dated the FO of G. pliozea at 5.6 Ma, the FO of G. puncticulata at 4.5 Ma, the LO of G. margaritae at 3.58 Ma, and the FO of G. inflata at 2.09 Ma. Although this mainly Mediterranean record may not be considered as a standard also for southern mid-latitudes, the sequence of these events may remain consistent should it not be disturbed by an incomplete local statigraphic section or by variations in the distribution of species. Consequently, we proposed a new zonation scheme, a hybrid of the existing subtropical and temperate zones. We hoped our scheme not only served a better biostratigraphy in this study but for the entire southern Australian region at large.
Figure F2 summarizes the integrated Neogene geochronostratigraphy, showing standard planktonic foraminiferal Neogene (N-) zones of Blow (1979) and Miocene (M-), Miocene transitional (Mt-), and Pliocene (PL-) zones and datum levels of Berggren et al. (1995). Jenkins' (1971, 1985, 1993) southern mid-latitude zones were correlated by recalculating the zonal markers. Stratigraphically useful datum levels found in southern Australia, as tabled in McGowran (1986) and McGowran et al. (1997b), were tied to the standard zonal scheme to highlight their synchronous nature. The new southern Australian Neogene SAN zones were recognized with species datums as detailed below, together with brief discussions on the regional assemblages.
Early Miocene Zones
Zone SAN1
Zone SAN1 (= N4a), 23.8-23.2 Ma, base = FO of Paragloborotalia kugleri sensu stricto (s.s.), top = LO of Globoquadrina dehiscens. The LCO of Turborotalia euapertura can be used as a proxy to the FO of P. kugleri in locating the Oligocene/Miocene boundary dated at 23.8 Ma by Berggren et al. (1995). Although Berggren (1992) reported the LO of T. euapertura coinciding the boundary in mid- to high-latitude sites, rare specimens of T. euapertura have been identified from the early Miocene of Australia (Li and McGowran, 2000; Li et al., 2000) and New Zealand (Jenkins, 1993). The common occurrence of Globigerinoides primordius can also be used to recognize the base of the Miocene. In southeastern Australia where P. kugleri and G. primordius are often absent, the LCO of T. euapertura may be more helpful for locating the base of Zone SAN1.
Zone SAN2
Zone SAN2 (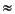 N4b), 23.2-21.6 Ma, base = FO of G. dehiscens (23.2 Ma), top = Globoconella incognita. However, G. dehiscens is rare in its early range and needs careful distinction from its predecessor G. praedehiscens. In New Zealand, these two morphotypes are often lumped as G. dehiscens, with an estimated FO as early as 25 Ma (Morgans et al., 1996). The FO of Globoturborotalita brazieri at ~24 Ma (Hornibrook et al., 1989) can also be used to justify the Oligocene/Miocene boundary, especially when accompanied by G. dehiscens, G. primordius, and Dentogloboqudrina globularis (Li et al., 1999). Within Zone SAN2, two other datums also occur: the FO of Globoturborotalita woodi and the FO of G. connecta. Ages estimated for these two datums range between 23 and 22.6 and 22 Ma and between 22.2 and 20.9 and 21 Ma by Hornibrook et al. (1989), Chaproniere et al. (1996), and Li and McGowran (2000), respectively. Li and McGowran (2000) also regarded the FO of G. connecta close to the Zone SAN2/SAN3 boundary at ~21.6 Ma, but this placement of the datum appears too high compared to that documented by Hornibrook et al. (1989). In this study, we placed the FO of F. connecta in upper Zone SAN2, at ~22.2 Ma, as did Carter, McCave, Richter, Carter, et al. (1999) (Fig. F2).
N4b), 23.2-21.6 Ma, base = FO of G. dehiscens (23.2 Ma), top = Globoconella incognita. However, G. dehiscens is rare in its early range and needs careful distinction from its predecessor G. praedehiscens. In New Zealand, these two morphotypes are often lumped as G. dehiscens, with an estimated FO as early as 25 Ma (Morgans et al., 1996). The FO of Globoturborotalita brazieri at ~24 Ma (Hornibrook et al., 1989) can also be used to justify the Oligocene/Miocene boundary, especially when accompanied by G. dehiscens, G. primordius, and Dentogloboqudrina globularis (Li et al., 1999). Within Zone SAN2, two other datums also occur: the FO of Globoturborotalita woodi and the FO of G. connecta. Ages estimated for these two datums range between 23 and 22.6 and 22 Ma and between 22.2 and 20.9 and 21 Ma by Hornibrook et al. (1989), Chaproniere et al. (1996), and Li and McGowran (2000), respectively. Li and McGowran (2000) also regarded the FO of G. connecta close to the Zone SAN2/SAN3 boundary at ~21.6 Ma, but this placement of the datum appears too high compared to that documented by Hornibrook et al. (1989). In this study, we placed the FO of F. connecta in upper Zone SAN2, at ~22.2 Ma, as did Carter, McCave, Richter, Carter, et al. (1999) (Fig. F2).
Zone SAN3
Zone SAN3 ( N5), 21.6-19 Ma, base = FO of G. incognita, top = FO of Globigerinoides trilobus sensu lato (s.l.). The zonal species G. incognita first appeared at 21.6 Ma (Berggren, 1992) and is mainly restricted to Zone N5 equivalents (= SAN3) in Australia (Li et al., 2000) and New Zealand (Hornibrook et al., 1989, fig. 27), even though it has been recorded as young as Zone N7 elsewhere (Kennett and Srinivasan, 1983; Berggren et al., 1995). Similar to the underlying Zone SAN2 assemblage, the cancellate-spinose G. woodi-G. connecta group is predominant but may be replaced in some intervals in relative abundance by the spinose Globigerina bulloides and microperforate tenuitellids that signal cooler and more fertile water (McGowran and Li, 1997; Li and McGowran, 2000).
Zone SAN4
Zone SAN4 (= uppermost N5 to lower N6), 19-18.5 Ma, base = FO of G. trilobus, top = FO of Globorotalia praescitula. These two datum levels appear to be diachronous, as Jenkins (1985, 1993) found the latter preceding the former and Hornibrook et al. (1989) reported their occurrences from a single level.
Zone SAN5
Zone SAN5 (= upper N6), 18.5-17.3 Ma, base = FO of G. praescitula, top = LO of Catapsydrax dissimilis. However, to separate this zone from the overlying Zone SAN6 is difficult because the marker species C. dissimilis is often rare and its datum is ambiguous locally. We discuss the characteristics of these two zonal assemblages together below.
Zone SAN6
Zone SAN6 (= N7), 17.3-16.4 Ma, base = LO of C. dissimilis, top = FO of Praeorbulina sicana. Both G. praescitula and Globoconella zealandica characterize the assemblage in Zones SAN5 and SAN6, although G. praescitula rarely ranges into the uppermost Zone SAN6. Other common species include G. trilobus, G. woodi, G. bulloides, and G. dehiscens. G. conoidea occurs sporadically within Zone SAN6. Subzones SAN6a and SAN6b (= Mt4a and Mt4b, respectively) can be recognized by the FO of Globoconella miozea at 16.7 Ma. A decline in the abundance of G. trilobus and G. woodi toward the top of Zone N7 is countered by an increase in G. bulloides in the Lakes Entrance section (Li and McGowran, 2000). Scott et al. (1990) reported from New Zealand that the FO of Fohsella peripheroronda was in the lower part of Zone N7, at ~17 Ma, but in southern Australia it often arrived much later, in Zone SAN8.
Middle Miocene Zones
Zone SAN7
Zone SAN7 (= N8a), 16.4-16.1 Ma, base = FO of P. sicana, top = FO of Praeorbulina glomerosa s.s. Apart from the marker species, both Globigerinoides (many species including G. trilobus and G. quadrilobatus) and Globoconella (mainly G. miozea, G. conica, and G. conoidea) are abundant in Zone SAN7 and range into younger intervals. Globigerina, however, declines to a minimum and is often represented by some large specimens of G. bulloides.
Zone SAN8
Zone SAN8 (= N8b), 16.1-15.1 Ma, base = FO of P. glomerosa s.s., top = FO of Orbulina suturalis. The assemblage is dominated by various species of Praeorbulina, Globigerinoides, and Globoconella.
Zone SAN9
Zone SAN9 (= N9), 15.1-14.8 Ma, base = FO of O. suturalis, top = LCO of P. glomerosa. Although many species are similar to those in Zone N8, the keeled globorotaliids began to become common in Zone N9, especially Globoconella panda, G. scitula, and the Menardella archeomenardii-Menardella praemenardii bioseries.
Zone SAN10
Zone SAN10 (= lower N10), 14.8-13.5 Ma, base = LCO of P. glomerosa, top = LO of F. peripheroronda. The age of 13.5 Ma for the latter datum is here proposed pending further studies, although it has been dated at 14.0 Ma by Berggren et al. (1995) and 13.0 Ma by Morgans et al. (1996). Other important events occurring within Zone SAN10 include the FO of M. praemenardii and the FO of Sphaeroidinellopsis kochi (both close to base of N10, according to Kennett and Srinivasan, 1983).
Zone SAN11
Zone SAN11 (= upper N10), 13.5-13 Ma, base = LO of F. peripheroronda, top = LO of Globoconella conica. Morgans et al. (1996) considered the later datum close to 13 Ma, as quoted also in Carter, McCave, Richter, Carter, et al. (1999). Two other datums are significant: the FO of Neogloboquadrina nympha at 13.4 Ma (Berggren, 1992) and the FCO of Paragloborotalia mayeri at 13.2 Ma (Scott et al., 1990). Jenkins (1993) recognized his Zone SN7 using the FO of P. mayeri, a datum he found from below the LO of F. peripheroronda.
Zone SAN12
Zone SAN12 ( N11-N13), 13-11.8 Ma, base = LO of G. conica, top = FO of Globoturborotalita nepenthes. Within this interval also occur the FOs of Menardella menardii, Globorotalia lenguaensis, and Globigerinatella siphonifera (all lower N12; ~12 Ma) (Kennett and Srinivasan, 1983) and the LO of Tenuitella minutissima (~12.3 Ma) (Li et al., 1992). P. mayeri is the dominant species.
Zone SAN13
Zone SAN13 (= N14), 11.8-11.4 Ma, base = FO of G. nepenthes, top = LO of P. mayeri. Scott et al. (1990) estimated the latter datum in New Zealand to be at 10.9 Ma, which is 0.5 Ma younger than that estimated by Berggren et al. (1995). The FO of Globorotalia scitula is close to the upper limit of Zone SAN13 at ~11.5 Ma. Abundant P. mayeri characterizes Zones SAN11 through SAN13 in an interval equivalent to Zone SN7 of Jenkins (1993).
Late Miocene Zones
Zone SAN14
Zone SAN14 (= N15), 11.4-10.9 Ma, base = LO of P. mayeri, top = FO of Neogloboquadrina acostaensis. According to Kennett and Srinivasan (1983), typical Globigerinoides ruber first occurred within this interval (~11 Ma).
Zone SAN15
Zone SAN15 (= lower N16), 10.9-9.2 Ma, base = FO of N. acostaensis, top = FCO of Neogloboquadrina pachyderma (9.2 Ma) (Berggren, 1992). Other important datums include the LO of G. panda (~10.3 Ma) (Scott et al., 1990) and a temporary LO of G. dehiscens (9.9 Ma) (Morgans et al., 1996).
Zone SAN16
Zone SAN16 (= upper N16), 9.2-8.3 Ma, base = FCO of N. pachyderma, top = FO of Globorotalia pleisiotumida or FO of Globigerinoides extremus. The LO of Neogloboquadrina continuosa was within this interval at ~8.5 Ma. G. conoidea-Globoconella miotumida complex remained common to abundant.
Zone SAN17
Zone SAN17 (= lower N17), 8.3-7.1 Ma, base = FO of G. pleisiotumida or FO of G. extremus, top = FO of Globoconella conomiozea. Note that the latter datum was dated at 6.9 Ma by Berggren et al. (1995), ~0.2 Ma younger than the age given in Morgans et al. (1996), which is adopted here. Common in Zones SAN17 and SAN18 include such keeled globorotaliids as M. menardii and G. conoidea-G. miotumida. The FOs of Globoconella juanai and Globoconella cibaoensis were from this interval, both at ~7.8 Ma.
Zone SAN18
Zone SAN18 (= mid N17, = upper Mt9), 7.1-6.0 Ma, base = FO of G. conomiozea, top = FO of G. margaritae or LO of G. lenguaensis.
Zone SAN19
Zone SAN19 (= upper N17), 6.0-5.3 Ma, base = FO of G. margaritae or LO of G. lenguaensis, top = LCO of Globoconella sphericomiozea. The FCO of Globoconella sphericomiozea or the FO of Globorotalia tumida (both at 5.6 Ma) can be used to separate between Subzones SAN19a and SAN19b, the latter being an analog to the Globorotalia sphericomiozea Subzone of Hornibrook et al. (1989). As in the underlying SAN18, Subzone SAN19a contains frequent M. menardii, Globigerinoides spp., Neogloboquadrina spp., Globorotalia plesiotumida, G. conoidea, G. juanai-G. cibaoensis complex, and G. miotumida. G. dehiscens last occurred within Subzone SAN19a, at ~5.8 Ma (Chaproniere et al., 1996). It occurs below the FCO of Globoconella sphericomiozea rather than concurs at 5.6 Ma, as reported by Berggren et al. (1995). Globoconella pliozea first appeared in Subzone SAN19b and ranged up to the Pliocene.
Pliocene Zones
Zone SAN20
Zone SAN20 ( N18, PL1a), 5.3-4.6 Ma, base = LCO of Globoconella sphericomiozea, top = LO of Globorotalia cibaoensis. Other common species include Globorotalia crassaformis, G. margaritae, Globoconella pliozea, and G. extremus.
Zone SAN21
Zone SAN21 ( N19, PL1b), 4.6-4.2 Ma, base = LO of Globorotalia cibaoensis, top = LO of G. nepenthes. Globoconella puncticulata first appeared close to the base of Zone SAN21 at ~4.5 Ma. Most common species are Globoconella puncticulata, G. crassaformis, Globoturborotalita spp., Globigerinoides spp., G. bulloides, and Globigerina falconensis.
Zone SAN22
Zone SAN22 ( N20, PL2), 4.2-3.7 Ma, base = LO of G. nepenthes, top = FO of Globoconella inflata at ~3.7 Ma (Morgans et al., 1996). As in the underlying Zone SAN21, G. crassaformis and Globoconella puncticulata are common to abundant.
Zone SAN23
Zone SAN23 (= lower N21, = PL3-PL4), 3.58-3.1 Ma, base = LO of G. margaritae, top = LO of Dentogloboquadrina altispira. The local assemblage is characterized also by abundant G. ruber and Globoconella inflata, as well as frequent G. bulloides, G. falconensis, and G. quinqueloba.
Zone SAN24
Zone SAN24 (upper N21, PL5-PL6), 3.1-2.0 Ma, base = LO of D. altispira, top = FO of Globorotalia truncatulinoides. The LCO of G. extremus was within Zone SAN24 at ~2.5 Ma, close to the FO of Globorotalia tosaensis. Species common to Zone SAN24 are Globoconella inflata, G. ruber, G. crassaformis, G. bulloides, and G. quinqueloba.
Zone SAN25
Zone SAN25 (= uppermost N21), 2.0-1.77 Ma, base = FO of G. truncatulinoides, top = LO of Globigerinoides obliquus or LO of G. woodi as a proxy. Apart from Globoconella inflata and G. ruber, G. crassaformis and Globigerina spp. are also common.
