Table
1 and Figure 1 provide
vertical profiles for total organic carbon (TOC) and C/N ratios (TOC/total
nitrogen, expressed by weight), total sulfur (total S), and ![]() 13C
of TOC in the sediment samples of Core 167-1017E-1H. TOC concentrations in Hole
1017E range from 4 to 31 mg/g (dry sediments). The C/N ratios range from 7.1 to
10.2. Significant differences in TOC concentrations, C/N values, and
13C
of TOC in the sediment samples of Core 167-1017E-1H. TOC concentrations in Hole
1017E range from 4 to 31 mg/g (dry sediments). The C/N ratios range from 7.1 to
10.2. Significant differences in TOC concentrations, C/N values, and ![]() 13C
of organic matter (OM) are seen between the Holocene (sections above 222.7 cm in
depth; <10.1 ka) and glacial period (sections below 222.7 cm in depth), as
summarized in Table 2. TOC and
total N concentrations in Holocene sections are roughly 1.8-1.9 times higher
than those in glacial sections. C/N values for the former (8.7) resemble those
for the latter (9.2). TOCs in Holocene sections are isotopically slightly
heavier than those in glacial ones.
13C
of organic matter (OM) are seen between the Holocene (sections above 222.7 cm in
depth; <10.1 ka) and glacial period (sections below 222.7 cm in depth), as
summarized in Table 2. TOC and
total N concentrations in Holocene sections are roughly 1.8-1.9 times higher
than those in glacial sections. C/N values for the former (8.7) resemble those
for the latter (9.2). TOCs in Holocene sections are isotopically slightly
heavier than those in glacial ones.
C/N ratios for OM of
marine origin generally range from 6 to 9 because of the high protein content of
lower organisms such as phytoplankton and zooplankton (e.g., Müller, 1977).
Higher plant-derived OM gives higher C/N values than that of marine origin
because it has a high percentage of nonprotein materials (e.g., 20-546; Meyers
and Ishiwatari, 1993). Thus, the present results of C/N for Hole 1017E may
indicate that OM is of marine origin. ![]() 13C
of TOC (
13C
of TOC (![]() 13CTOC)
supports this conclusion because the
13CTOC)
supports this conclusion because the ![]() 13CTOC
values for Core 167-1017E-1H range from -21.0
13CTOC
values for Core 167-1017E-1H range from -21.0![]() to -22.8
to -22.8 ![]() , which are
common to the organic matter of marine origin. A similar conclusion was reached
by Gardner et al. (1997), who found
, which are
common to the organic matter of marine origin. A similar conclusion was reached
by Gardner et al. (1997), who found ![]() 13CTOC
in the sediment cores from a similar region (35°N,
121°W) roughly -22
13CTOC
in the sediment cores from a similar region (35°N,
121°W) roughly -22![]() over the last 60 k.y. It is noteworthy that
over the last 60 k.y. It is noteworthy that ![]() 13CTOC
values in the Holocene are ~0.8
13CTOC
values in the Holocene are ~0.8![]() higher than those in glacial sections, and
higher than those in glacial sections, and ![]() 13CTOC
values in glacial are relatively constant. The invariability of
13CTOC
values in glacial are relatively constant. The invariability of ![]() 13CTOC
values in glacial contrasts with the synchronous fluctuations in TOC, C/N, and
total S. Most of the sections with low TOC concentrations, C/N, and total S
concentrations in glacial are sandy (fine or coarse grained) deposits (~320,
~360, ~380, ~395, ~420, ~440, ~460, ~490, and ~507 cm in depth, respectively; Fig.
1). The stratigraphy of the sediment core is given by Tada et al. (Chap.
25, this volume). These sections are probably well oxygenated, which
coincides with low total S contents. Low C/N ratios in the sandy sections may be
produced as a result of contribution of inorganic ammonium sorbed by clays to
total nitrogen, which often becomes important in TOC-poor sediment (Müller,
1977). A marked positive shift in
13CTOC
values in glacial contrasts with the synchronous fluctuations in TOC, C/N, and
total S. Most of the sections with low TOC concentrations, C/N, and total S
concentrations in glacial are sandy (fine or coarse grained) deposits (~320,
~360, ~380, ~395, ~420, ~440, ~460, ~490, and ~507 cm in depth, respectively; Fig.
1). The stratigraphy of the sediment core is given by Tada et al. (Chap.
25, this volume). These sections are probably well oxygenated, which
coincides with low total S contents. Low C/N ratios in the sandy sections may be
produced as a result of contribution of inorganic ammonium sorbed by clays to
total nitrogen, which often becomes important in TOC-poor sediment (Müller,
1977). A marked positive shift in ![]() 13CTOC
values around 10 ka can be explained as follows. Carbon isotopic composition of
TOC in sediment is generally controlled by the following three factors: the
nature of source material (terrestrial higher plants vs. marine phytoplankton),
temperatures of the photosynthetic fixation of CO2, and rate of
photosynthesis (if we assume that TOC in sediment is derived from marine
phytoplankton alone). Following Schoell et al. (1994), the relationship among
the three factors can be expressed as
13CTOC
values around 10 ka can be explained as follows. Carbon isotopic composition of
TOC in sediment is generally controlled by the following three factors: the
nature of source material (terrestrial higher plants vs. marine phytoplankton),
temperatures of the photosynthetic fixation of CO2, and rate of
photosynthesis (if we assume that TOC in sediment is derived from marine
phytoplankton alone). Following Schoell et al. (1994), the relationship among
the three factors can be expressed as
where ![]() 13Ci
is the carbon isotopic composition of TOC in sediment, obtained after correction
for a shift in
13Ci
is the carbon isotopic composition of TOC in sediment, obtained after correction
for a shift in ![]() 13C
resulting from early diagenesis in sediments (Hayes et al., 1983);
13C
resulting from early diagenesis in sediments (Hayes et al., 1983); ![]() 13CDIC
is the isotopic composition of the dissolved CO2; and
13CDIC
is the isotopic composition of the dissolved CO2; and ![]() p
is the fractionation between dissolved CO2 and primary photosynthate,
as shown above.
p
is the fractionation between dissolved CO2 and primary photosynthate,
as shown above. ![]() p
is written as
p
is written as
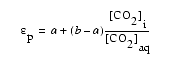 , (2)
, (2)
where a
and b are the
fractionations associated with diffusion of CO2 into the
phytoplankton (0.7![]() ) and
fixation of CO2 with the phytoplankton (27.0
) and
fixation of CO2 with the phytoplankton (27.0![]() ;
Yoshioka, 1997). [CO2]i is intracellular CO2
concentration, and [CO2]aq is extracellular concentration.
The ratio [CO2]i /[CO2]aq is assumed
to be photosynthetic carbon demand and should be related to the phytoplankton
growth rate when transport of CO2 into the cell is controlled by
diffusion, which is common in marine systems (Shemesh et al., 1993; Laws et al.,
1995).
;
Yoshioka, 1997). [CO2]i is intracellular CO2
concentration, and [CO2]aq is extracellular concentration.
The ratio [CO2]i /[CO2]aq is assumed
to be photosynthetic carbon demand and should be related to the phytoplankton
growth rate when transport of CO2 into the cell is controlled by
diffusion, which is common in marine systems (Shemesh et al., 1993; Laws et al.,
1995). ![]() 13CDIC
can be estimated from
13CDIC
can be estimated from ![]() 13C
of carbonate tests of planktonic foraminifers and paleo-sea-surface temperatures
(paleo-SSTs) by using the following empirical equation (Jasper et al., 1994):
13C
of carbonate tests of planktonic foraminifers and paleo-sea-surface temperatures
(paleo-SSTs) by using the following empirical equation (Jasper et al., 1994):
where ![]() t
presents the isotopic composition of foraminifera calcite and T is paleo-SST (in
Kelvin).
t
presents the isotopic composition of foraminifera calcite and T is paleo-SST (in
Kelvin).
The difference in ![]() 13C
for sedimentary TOC at two different times (
13C
for sedimentary TOC at two different times (![]() 13Ci)
can be roughly expressed as
13Ci)
can be roughly expressed as
where ![]() 13CDIC
is the difference in
13CDIC
is the difference in ![]() 13C
of dissolved CO2 in seawater, which is estimated from the difference
in
13C
of dissolved CO2 in seawater, which is estimated from the difference
in ![]() 13C
of carbonate tests of planktonic foraminifers deposited at the different times
with correction of paleo-SST.
13C
of carbonate tests of planktonic foraminifers deposited at the different times
with correction of paleo-SST.
The term ![]()
![]() p,
which is related to the difference in photosynthetic carbon demand at different
times, can be calculated from Equation 4. The term
p,
which is related to the difference in photosynthetic carbon demand at different
times, can be calculated from Equation 4. The term ![]()
![]() p
was named as a parameter related to comparative growth rate (PCGR; Ishiwatari et
al., 1999). A high PCGR at a particular time indicates a low photosynthetic
carbon demand (i.e., growth rate of phytoplankton would be low) relative to that
at the reference time.
p
was named as a parameter related to comparative growth rate (PCGR; Ishiwatari et
al., 1999). A high PCGR at a particular time indicates a low photosynthetic
carbon demand (i.e., growth rate of phytoplankton would be low) relative to that
at the reference time.
According to Cannariato (pers.
comm., 1998), ![]() 13C
of inorganic carbon (IC) of planktonic foraminifera (Globigerina
bulloides) tests ranges roughly from -1.4
13C
of inorganic carbon (IC) of planktonic foraminifera (Globigerina
bulloides) tests ranges roughly from -1.4![]() to 0.6
to 0.6![]() in 280-160 cm in
depth (13-7 ka) and shows no marked trend. Sea-surface temperature (SST)
estimated from alkenone analysis indicates a rapid increase from ~12°C
at 230 cm in depth (10.5 ka) to ~15°C
at ~210 cm in depth (9.5 ka; R. Ishiwatari et al., unpubl. data. In addition,
in 280-160 cm in
depth (13-7 ka) and shows no marked trend. Sea-surface temperature (SST)
estimated from alkenone analysis indicates a rapid increase from ~12°C
at 230 cm in depth (10.5 ka) to ~15°C
at ~210 cm in depth (9.5 ka; R. Ishiwatari et al., unpubl. data. In addition, ![]() 13C
values of IC of planktonic foraminifera tests fluctuate around -1
13C
values of IC of planktonic foraminifera tests fluctuate around -1![]() and show no significant difference before and after 10 ka. Thus,
and show no significant difference before and after 10 ka. Thus, ![]() 13C
of the dissolved CO2 is estimated to become higher by ~0.3
13C
of the dissolved CO2 is estimated to become higher by ~0.3![]() (
(![]() 13CDIC
= ~0.3
13CDIC
= ~0.3![]() ) at the SST
transition at ~10 ka. Consequently,
) at the SST
transition at ~10 ka. Consequently, ![]()
![]() p
equals ~-0.7
p
equals ~-0.7![]() . This
means that the increase in growth rate of phytoplankton, which may correspond
closely to planktonic primary productivity, might be responsible for the ~0.5
. This
means that the increase in growth rate of phytoplankton, which may correspond
closely to planktonic primary productivity, might be responsible for the ~0.5![]() positive shift in
positive shift in ![]() 13C
values of sedimentary TOC at ~10 ka, although a substantial range of
possibilities must be considered.
13C
values of sedimentary TOC at ~10 ka, although a substantial range of
possibilities must be considered.
Another factor that can
cause a part of the positive shift in ![]() 13CTOC
values around 10 ka is a drop in input of terrestrial higher plant-derived OM,
which is suggested by lignin analysis (R. Ishiwatari et al., unpubl. data) of
the sediment core. The lignin analysis indicates that TOC-normalized
concentrations of lignin phenols in Holocene sediments are one-half to
one-quarter times lower than those in glacial. However, we cannot evaluate at
present how the drop in input of terrestrial higher plant-derived OM contributes
to the positive shift in the
13CTOC
values around 10 ka is a drop in input of terrestrial higher plant-derived OM,
which is suggested by lignin analysis (R. Ishiwatari et al., unpubl. data) of
the sediment core. The lignin analysis indicates that TOC-normalized
concentrations of lignin phenols in Holocene sediments are one-half to
one-quarter times lower than those in glacial. However, we cannot evaluate at
present how the drop in input of terrestrial higher plant-derived OM contributes
to the positive shift in the ![]() 13CTOC
value.
13CTOC
value.
Sulfur concentrations are high in sections in the 306-280 cm range (25.3-13.1 ka), which matches excellently with anoxia Event 1, corresponding to the number 1 (Bølling/Allerød) of warm interstadials (15.5-12.7 ka; Behl and Kennett, 1996). Behl and Kennett (1996) evaluated the degree of oxygenation of sediments in the central Santa Barbara Basin (ODP Hole 893A: 34°17.25´N, 120°2.2´W: 576.5 m water depth), which is close to Site 1017, using the bioturbation index. An oxygenation level for the anoxia event indicates anaerobic/dysaerobic boundary conditions allowing only micro- to meiofaunal bioturbation sufficient to diffuse but not destroy primary laminations (~0.1 mL O2/L; Behl and Kennett, 1996). The marked correlation of the high sulfur section to anoxia Event 1 suggests that the same factor (the presence of relatively old bottom waters) played a role for both events. This result seems to support the idea of Behl and Kennett (1996) on the bottom-water system in the California margin.
The section corresponding to anoxia Event 2 at 23.7-22.8 ka reported by Behl and Kennett (1996) does not show high sulfur content, probably because the oxygenation level (>0.3 mL O2/L dysaerobic to aerobic) is not enough for producing high-sulfur sediments.
High-sulfur contents are observed for other sections (400 cm and 455-430 cm in depth). But the corresponding low benthic oxygenation was not seen in the Santa Barbara Basin (Behl and Kennett, 1996). These results suggest that short-term changes in the oxygenation level occur locally during glacial periods.
![]()