BIOSTRATIGRAPHY
Cenozoic
Miocene planktonic foraminifers are rare and only one sample yielded age information. The presence of Globorotalia praescitula (first appearance = 18.8 Ma), Sphaeroidinella disjuncta (Zones N6–N11; ~18.8–12.5 Ma), and Globorotalia zealandica (approximately Zones N6–N8; ~18.8–15.2 Ma) at 86.0 ft (26.21 m) (Fig. F2) suggests an age less than ~18.8 Ma.
Eocene planktonic foraminifers are rare and are not generally well preserved (Table T3). Because of the nearshore location of the Sea Girt corehole, marker species are rare and zonation has relied upon secondary markers.
A single specimen assigned to the genus Truncorotaloides is found in the Shark River Formation at 176 ft (53.64 m) indicating the sediments below are not younger than Biochron P14 (Fig. F2). Orbulinoides beckmanni is rarely found in New Jersey, thus Zones P12–P14 cannot be subdivided. The highest occurrence of Morozovella aragonensis (the marker for the base of Zone P12) at 276 ft (84.12 m) is believed to be premature based on comparisons with nannofossil biostratigraphy (Figs. F2, F10). The first occurrence of Turborotalia pomeroli at 235 ft (71.63 m) indicates that the sediments above the sequence boundary at 236.1 ft (71.96 m) should be assigned to Zone P12 or younger (Berggren et al., 1995). Globigerinatheka subconglobata is first found at 251 ft (76.50 m) and is used to approximate the Zone P11/P10 boundary. If the basal Zone P11 assignment is correct, then the sequence from 236.1 to 265.8 ft (71.96 to 81.02 m) straddles the Zone P10/P11 boundary. Most New Jersey cores contain an expanded sequence that straddles the Zone P11/P12 boundary (Sequence E7). Calcareous nannofossils cannot resolve whether these sediments should be assigned to Subzone NP15b or NP15c. If the sediments can be assigned to calcareous nannofossil Subzone NP15b, then this sequence should be assigned to Sequence E6a previously known only from the Ancora corehole and Sequence E7 is missing.
Zones P9 and P10 cannot be distinguished in the cores because of the absence of Hantkenina. The base of Zone P9 is approximated at 266 ft (81.08 m) by the first occurrence of Acarinina bullbrooki (Fig. F3). The base of Zone P8 is approximated at 290.4 ft (88.51 m) by the first occurrence of Subbotina inaequispira (Fig. F3). The first occurrence of M. aragonensis, at 316 ft (96.32 m), defines the base of Zone P7 (Fig. F3). The section between 316 and 341 ft (96.32 and 103.94 m) is difficult to zone because of the absence of Morozovella formosa in the core. The presence of Morozovella gracilis and Morozovella subbotinae in this interval is typical of a Subzone P6b fauna. The assignment to calcareous nannoplankton Zone NP11 also suggests that these sediments correlate to Subzone P6b. Lowermost Eocene sediments from 346 to 361 ft (105.46 to 110.03 m) are barren of foraminifers.
Paleocene planktonic foraminifers are rare and generally poorly preserved, making a detailed zonation impossible. A sample at 366 ft (111.56 m) contains Acarinina primitiva, but members of the Paleocene/Eocene boundary excursion fauna are absent. Samples from 376 to 446 ft (114.60 to 135.94 m) are barren of foraminifers. A sample at 451 ft (137.46 m) contains a few benthic foraminifers and a single specimen of Subbotina triangularis. Samples at 456 and 461 ft (138.99 and 140.51 m) contain Globanomalina pseudomenardii, indicating assignment to Zone P4 (Fig. F3). The sample at 461 ft (140.51 m) contains Acarinina subsphaerica indicating assignment to Subzone P4a (Fig. F3). The absence of A. subsphaerica from the sample at 456 ft (138.99 m) may indicate assignment to Subzone P4b. A sample at 457.2 ft is assigned to Subzone P3b through Zone P4, with Acarinina mckannai, A. subsphaerica, S. triangularis, Igorina pusilla, Morozovella acuta, Morozovella aequa, and Subbotina velascoensis. A sample at 476 ft (145.08 m) contains Morozovella angulata without Igorina albeari, perhaps indicating assignment to Subzone P3a, although the sample contains few planktonics. This assignment is consistent with calcareous nannofossil Subzone NP4b. Samples at 491 and 494 ft (149.66 and 150.57 m) contain Globanomalina compressa and Globoconusa daubjergensis, whose highest occurrence is in Subzone P1c (Olsson et al., 1999) and indicates assignment to Subzone P1c. A sample from 495.2 ft contains a Danian assemblage including G. daubjergensis and Praemurica pseudoinconstans (Zone P1, probably Subzone P1a) as did a sample from 504.4 ft.
Cretaceous/Paleogene Boundary
Samples from 505.0 to 512.0 ft (153.96 to 156.09 m) were examined for planktonic foraminifers to identify the Cretaceous/Paleogene (K/Pg) boundary (Table T4). Planktonic foraminifers in this interval were relatively sparse, probably because of a shallow inner shelf paleodepth. The K/Pg boundary is placed between samples at 509.0 and 509.5 ft (155.18 and 155.34 m). The Danian interval from 509.0 ft (155.14 m) is placed in Subzone P1a based on the presence of Eoglobigerina edita, Eoglobigerina eobulloides, P. pseudoinconstans, Praemurica taurica, G. daubjergensis, Woodringina claytonensis, and Woodringina hornerstownensis. The zonal marker for Subzone P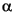 , Parvularugoglobigerina eugubina, was not identified. This may be because of the scarcity of planktonic foraminifers in the Danian interval. In other ODP New Jersey coastal plain coreholes, the species is rare, so its absence at Sea Girt is not surprising and is probably because of the shallow paleodepth. Likewise, without this species, Zone P0 cannot be identified.
, Parvularugoglobigerina eugubina, was not identified. This may be because of the scarcity of planktonic foraminifers in the Danian interval. In other ODP New Jersey coastal plain coreholes, the species is rare, so its absence at Sea Girt is not surprising and is probably because of the shallow paleodepth. Likewise, without this species, Zone P0 cannot be identified.
Cretaceous samples from 509.5 to 512.0 ft (155.34 to 156.09 m) contain rare specimens of Globotruncana, Rugoglobigerina, and Racemiguembelina, along with Guembelitria cretacea, which also occurs in the Danian interval. Other than indicating a Maastrichtian age, zonal markers are absent because of a shallow paleodepth. The presence of Pseudoguembelina hariaensis just below this interval, however, indicates uppermost Maastrichtian for this interval.
Maastrichtian
The Maastrichtian occurs from samples 509.5 to 560.0 ft (155.34 to 170.73 m), which are placed largely in the Navesink Formation except for the top 1 ft (0.3 m), which is placed in the Hornerstown Formation (Table T4). The base of the Navesink Formation is placed at 566.1 ft (172.59 m). The highest occurrences (HOs) of Globotruncana linneiana at 550 ft (167.64 m) and Rosita fornicata identify the top of the lower Maastrichtian Globotruncana aegyptiaca Zone. The upper Maastrichtian marker species Gansserina gansseri was not identified at Sea Girt, probably because of shallow paleodepth. The entire Maastrichtian section occurs within the Navesink sequence, which extends into the basal Danian with a shallowing upward HST across the K/Pg boundary (Olsson et al., 2002).
Campanian
From 570.4 to 835.0 ft (179.90 to 254.57 m), 28 samples were examined in the Mount Laurel, Wenonah, Marshalltown, and upper Englishtown Formations for planktonic foraminifers (Tables T5). Samples were taken every 5 ft (1.5 m) in the transgressive Marshalltown Formation and usually every 10 ft (3.0 m) in the regressive highstand formations, where recovery of foraminifers was expected to be sparse or none.
Planktonic foraminifers were few and very rare in the Mount Laurel, Wenonah, and Marshalltown Formations. Except for one sample, the upper Englishtown Formation was barren of planktonic foraminifers (Table T5). None of the recovered species were zonal markers.
Turonian–Cenomanian
The Bass River Formation (1290.7–1495.0 ft; 393.51–455.79 m) was sampled from 1346.0 to 1477.0 ft (410.36 to 450.30 m), where foraminifers were first noted; the upper section is barren. Planktonic foraminifers (Table T6) were abundant in only one sample (1396.0 ft; 425.61 m). The most common species identified in the formation were Guembelitria cenomana and Whiteinella archeocretacea. Foraminifers were sparse or absent in most samples and diversity was very low, reflecting the shallow inner shelf paleoenvironment. The interval from 1456.0 to 1500.0 (443.90 to 457.32 m) was barren, as was sample 1431.0 ft (436.28 m). The Cenomanian/Turonian boundary, based on planktonic foraminifers, could not be identified because Rotalipora did not occur, again reflecting the shallow paleodepth.
Cenozoic
Benthic foraminifers from the Eocene are generally abundant (Table T7), well preserved, and record environments ranging from middle to outer neritic paleodepths. The exceptions are from 140 to 171 ft (42.67 to 52.12 m), which is barren, and from 346 to 365 ft (111.25 to 105.46 m), which contains very few, poorly preserved specimens. Upper middle Eocene Sequence E8 (140–236.1 ft; 42.67–71.96 m) contains an assemblage including Cibicidina, Globobulimina, Guttulina, Gyroidinoides, Lenticulina, and Pararotalia. The sample at the base of the sequence (235 ft; 71.63 m) is dominated by Cibicidoides aff. subspiratus and was deposited at paleodepths of ~100 m (Biofacies C of Browning et al., 1997). The remainder of the sequence is dominated by species of Cibicidina, Globobulimina, and Pararotalia, indicating paleodepths of ~50 m (Biofacies A of Browning et al., 1997).
Sequence E7 (235–265 ft; 71.63–80.77 m) is dominated by species of Alabamina, Cibicidoides, Guttulina, Gyroidinoides, Lenticulina, and Stilostomella. Samples at the base of the sequence (251–261 ft; 76.70–79.55 m) are dominated by C. subspiratus, indicating paleodepths of ~135 m (Biofacies G of Browning et al., 1997). Samples at the top of the sequence (241–246 ft; 73.46–74.98 m) are dominated by Alabamina and Hanzawaia, indicating paleodepths of ~75 m (Biofacies B of Browning et al., 1997). One sample was examined from Sequence E5 (265–271 ft; 80.77–82.60 m). It contains C. subspiratus, Eponides, and Lenticulina, indicating paleodepths of ~135 m (Biofacies G of Browning et al., 1997).
Sequences in the Manasquan Formation contain the deepest paleowater depths recovered in the Eocene. Sequence E3 (271–318 ft; 82.60–96.93 m) contains four different biofacies that record deepening in the TST and slow shallowing in the HST. The sample at 316 ft (96.32 m) is dominated by Cibicidoides aff. pseudoungerianus, indicating paleodepths of ~100 m (Biofacies C of Browning et al., 1997). Samples from 290 to 311 ft (88.39 to 94.79 m) are dominated by C. aff. subspiratus indicating paleodepths of ~155 m (Biofacies E of Browning et al., 1997). A sample at 286 ft (87.17 m) is dominated by Siphonina claibornensis, indicating paleodepths of ~125 m (Biofacies D of Browning et al., 1997). Samples at the top of the sequence (271–281 ft; 82.60–85.65 m) are dominated by C. aff. pseudoungerianus, indicating shallowing to ~100 m. (Biofacies C of Browning et al., 1997). Sequence E2 (318–359 ft; 96.93–109.42 m) contains Gavelinella capitata and Cibicidoides eocaenus in samples from 326 to 341 ft (99.36 to 103.94 m), indicating paleodepths of ~185 m (Biofacies F of Browning et al., 1997). The sample at 321 ft (97.84 m), the top of the sequence, is dominated by C. aff. subspiratus, indicating paleodepths of ~155 m (Biofacies E of Browning et al., 1997).
Paleocene sequences generally contain few fossiliferous samples. Three samples (366, 371, and 376 ft; 111.56, 113.08, and 114.60 m) from the upper Paleocene–lowermost Eocene unnamed clay contain a fauna dominated by Pulsiphonina prima, indicating middle neritic paleodepths. Samples in the Vincentown Formation between 386 and 446 ft (117.65 and 135.94 m) are barren. A sample at 456 ft (138.99 m), near the base of Sequence Pa3, contains a fauna that includes Tappanina selmensis, indicating an outer neritic paleoenvironment. A sample (466 ft; 142.04 m) from Sequence Pa2 is dominated by P. prima but also contains T. selmensis, indicating middle to outer neritic paleodepths. Sequences Pa1a and Pa1b at the base of the Hornerstown Formation contain a general midway benthic foraminiferal fauna, indicating middle neritic paleodepths.
Maastrichtian
Benthic foraminiferal Biofacies 1–4 were first established in the Navesink sequence at the Bass River corehole. They are a useful means of correlation and establishing whether the Maastrichtian section is complete. These biofacies can be recognized in the Sea Girt corehole, allowing correlation with the other New Jersey coreholes. The occurrence of benthic foraminiferal species is shown in Table T8. Biofacies 2, which consistently occurs in all New Jersey coreholes at the top of the Maastrichtian and extends into the basal Danian, is characterized by the occurrence of Alabamina midwayensis and Anomalinoides acuta. This indicates that the topmost part of the Maastrichtian is present at the Sea Girt corehole despite the absence of planktonic foraminiferal markers. The downhole appearance of several biofacies species and planktonic zonal species is a useful correlation tool among New Jersey coreholes. Correlation of the Sea Girt corehole with other coreholes is shown in Figure F9.
Campanian
Benthic foraminifers were common in the Mount Laurel, Wenonah, and Marshalltown Formations. They ranged from numerous in some samples to few in other samples (Table T9). The upper Englishtown Formation varied from intervals that were barren to intervals with a scattering of foraminifers, reflecting the inner shelf to shoreface paleoenvironment. The most common species is Lenticulina pseudosecans, which was also present in samples of the Marshalltown Formation. Foraminifers were most abundant in the basal part of the Marshalltown Formation (676.0–681.0 ft; 206.10–207.62 m), reflecting the initiation of a transgressive cycle. The rest of the assemblages in the Mount Laurel Formation indicate a shallowing HST deposited in inner shelf paleodepths. Foraminifers were most abundant in the top sample of the Mount Laurel Formation, probably representing bioturbation and mixing with the overlying transgressive Navesink Formation.
Cenomanian–Turonian
Benthic foraminifers (Table T10) range from numerous to very rare in the Cenomanian–Turonian Bass River Formation. The lower interval from sample 1460.0 ft (445.12 m) was barren. Species of Epistomina were most useful in establishing correlation with the Bass River corehole and in placing the Cenomanian/Turonian boundary. The samples in the Turonian interval contain Epistomina chapmani and Epistomina stelligera, placing this interval in the lowermost Turonian below the interval with Epistomina sliteri and Epistomina lenticularia in the Bass River corehole. The Cenomanian/Turonian boundary is placed between samples 1415.0 ft (431.40 m) and 1421.0 ft (433.23 m) on the first occurrence of Epistomina suturalis. The diagnostic Cenomanian species G. cenomana was identified in sample 1445.0 ft (440.55 m), where it is common. The benthic foraminiferal assemblage of the Bass River Formation indicates a shallow inner shelf paleodepth of ~30 m or less.
Cenozoic
A total of 75 samples, taken at regular intervals of ~5 ft (1.52 m) between 141 and 508.5 ft (42.98 and 154.99 m), were analyzed for calcareous nannofossil biozonal interpretation. Examination with a Zeiss standard microscope at magnifications of x600 and x1200 was conducted on smear slides prepared directly from small amounts of sediment. A species inventory was attempted for every assemblage, but no quantitative record was established. Instead, attention was given to recovering markers species and determining their lowest occurrences (LOs) and/or HOs (Table T11). The biozonal framework used is that of Martini (1971) and Martini and Müller (1986).
In general, calcareous nannofossil abundance varies from common to rare throughout the section (Table T11). However, two intervals were barren: from 141 to 166 ft (42.98 to 50.60 m) and 401 to 446 ft (122.22 to 135.94 m). In addition, the lowermost 3.5 ft (1.07 m) of the section yielded extremely rare calcareous nannofossils. Diversity is low at most levels, but remarkably high in the lower middle Eocene (266 and 271 ft; 81.08 and 82.60 m; Subzone NP14a). Assemblages are generally rich in representatives of the genera Helicosphaera, Pontosphaera, and Rhabdosphaera, possibly reflecting coastal proximity. In contrast, they are generally poor in discoasters, except in the lower middle and upper lower Eocene (266–316 ft; 81.08–96.32 m; Subzone NP14a and Zone NP12). Notable are the absence of Discoaster mohleri (marker base of Zone NP7) and the scarcity of Sphenolithus furcatolithoides (a common marker of the Zones NP15–NP16 interval), Chiasmolithus grandis (whose HO marks the top of Zone NP17), Discoaster barbadiensis, Discoaster saipanensis, and Ericsonia formosa, Eocene taxa generally common at middle latitudes.
The 367-ft-thick (111.86 m) section examined is Paleogene, spanning the middle Eocene (Bartonian; Zone NP17) to lowermost Paleocene (lower Danian; Zone NP1). It is highly discontinuous, however, with most zonal boundaries occurring at unconformable contacts.
The youngest datable level (171 ft; 52.12 m) belongs to Zone NP17, as indicated by the scarce occurrence of C. grandis and Sphenolithus celsus and the absence of Chiasmolithus oamaruensis, the LO of which defines the base of Zone NP18. This co-occurrence was observed in the Barton beds of England (Aubry, 1983, 1985). Zone NP17 extends to 186 ft (56.69 m) and is characterized by the presence of Chiasmolithus expansus, Corannulus germanicus, Cruciplacolithus delus, Helicosphaera wilcoxonii, Reticulofenestra reticulata, Reticulofenestra umbilicus, and Neococcolithes minutus.
Samples at 191 and 196 ft (58.22 and 59.74 m) have yielded essentially uncharacteristic assemblages. A few placoliths of Chiasmolithus solitus were recovered at 196 ft (59.79 m). On this basis, 196 ft (59.79 m) is assigned to Zone NP16 and 191 ft (58.22 m) to Zone NP17.
The interval from 201 to 235 ft (61.26 to 71.35 m) belongs to Zone NP16. The assemblages include, among others, C. solitus (common except at 201 ft [61.26 m]), C. grandis, C. germanicus, Daktylethra punctulata, Discoaster barbadiensis (rare), Discoaster tanii nodifer (rare), Helicosphaera seminulum, Pontosphaera wechesensis, and occasionally S. furcatolithoides (206 ft; 62.79 m). The lowest occurrence of R. reticulata at 226 ft (68.88 m) marks the upper part of the zone.
There is a sharp difference in the composition of the assemblages at 235 ft and 241.5 ft (71.63 and 73.61 m). R. umbilicus is common and large at 235 ft (71.63 m) and absent at 241.5 ft (73.61 m). Both Nannotetrina sp. (overgrown) and Chiasmolithus gigas occur at this latter level, characterizing Subzone NP15b. Blackites gladius, the HO of which marks the top of Zone NP15, was not encountered, nor was Discoaster bifax, a species often used as a substitute marker for the base of Zone NP16. However, the frequency and size of R. umbilicus at 235 ft (71.63 m) is indicative of Zone NP16. C. gigas and overgrown specimens of Nannotetrina (remarkably abundant at 261 ft [79.55 m]) occur consistently between 241.5 and 261 ft (73.61 and 79.55 m), an interval thus assigned to Subzone NP15b.
Samples from 266 and 271 ft (81.08 and 82.60 m) are well characterized by abundant, well preserved, and diverse assemblages. These include C. solitus, C. expansus, C. grandis, D. barbadiensis, Discoaster boulangeri, Discoaster distinctus, Discoaster mirus, Discoaster nonaradiatus, Discoaster septemradiatus, Discoaster wemmelensis, Helicosphaera lophota, Helicosphaera seminulum, Lophodolithus mochlophorus, Lophodolithus nascens, and Trochastrites hohnensis. The occurrence of Discoaster sublodoensis, with common Discoaster lodoensis and rare (at 271 ft) Discoaster kuepperi, characterizes lower middle Eocene Subzone NP14a. Tribrachiatus orthostylus and D. lodoensis co-occur in the 276–316 ft (84.12–96.32 m) interval that is therefore assigned to Zone NP12.
There is an abrupt change in assemblage composition between the sample at 316 ft (96.32 m), where discoasters, among which D. lodoensis are common, and the sample at 321 ft (97.84 m), where discoasters are rare and there was no D. lodoensis. The absence of D. lodoensis at 321 ft (97.84 m) is not taken to reflect the scarcity of discoasters, because similar assemblages occur from this level to 356 ft (108.51 m). In this interval, T. orthostylus is common, associated with Discoaster binodosus and D. kuepperi, a trilogy characteristic of Zone NP11. The occurrences of Discoaster pacificus (scarce), Discoaster multiradiatus, and Ellipsolithus macellus support this zonal assignment.
The interval from 366 to 386 ft (111.56 to 117.65 m) yields rare to very rare calcareous nannofossils and is tentatively assigned to Zone NP9, based on the occurrence of Discoaster perpolitus, a species very closely related to D. multiradiatus (Romein, 1979). We note the absence of Fasciculithus spp. in this interval and the occurrence of Discoaster anartios at 376 ft (114.60 m), the occurrence of which marks the base of Subzone NP9b and with a range that is restricted to the stratigraphic extent of the carbon isotope excursion (Kahn and Aubry, 2004).
The HO of Fasciculithus tympaniformis lies at 391.5 ft (119.33 m). The interval between this level and 396 ft (120.70 m), in which D. perpolitus and F. tympaniformis co-occur, belongs to Subzone NP9a. Heliolithus riedeli is common at 451 and 460 ft (137.46 and 140.21 m), both assigned to Zone NP8. In the absence of H. riedeli and the presence of Heliolithus kleinpelli and Discoaster helianthus, 461 ft (140.51 m) is tentatively assigned to Zone NP7. Discoaster mohleri was not encountered at Sea Girt. H. kleinpelli is abundant at 466 ft (142.04 m), which belongs to Zone NP6. Samples at 471 and 476 ft (143.56 and 145.08 m) yield E. macellus, Fasciculithus billi, Fasciculithus janii, and Sphenolithus primus. In the absence of F. tympaniformis, they are assigned to Subzone NP4b. The LO of E. macellus lies at 494.7 ft (150.78 m), marking the base of Zone NP4. Consequently, the interval between this level and 481 ft (146.61 m) is assigned to Subzone NP4a. Cruciplacolithus tenuis is rare at 501 ft (152.70 m), whose assemblages include Cyclagelopshaera alta, Cyclagelopshaera margereli, Placozygus sigmoides, and Cretaceous species. Samples from 505 and 507 ft (153.92 and 154.53 m) are essentially barren. Markalius inversus and Cyclagelosphaera occur at 508 ft (154.84 m), without Cruciplacolithus primus. This level, with extremely rare coccoliths, probably belongs in the lower Danian Subzone NP1a.
Based on calcareous nannofossil stratigraphy alone, several unconformities are inferred in the sedimentary succession recovered from the Sea Girt corehole. The youngest unconformity is between 235 and 241.5 ft (71.63 and 73.61 m; lower Zone NP16 and Subzone NP15b). As mentioned above, the contrast in the composition of the assemblages at these levels is indicative of a stratigraphic gap, which encompasses all of Subzone NP15c. The next two unconformities occur between 261 and 266 ft (79.55 and 81.08 m; Subzones NP15a and NP14a) and between 271 and 276 ft (82.60 and 84.12 m; Subzones NP14a and Zone NP12) and belong to the set of unconformities associated with the lower/middle Eocene boundary on the western margin of the North Atlantic (Aubry, 1995).
The sharp change in assemblage composition between 316 and 321 ft (96.32 and 97.84 m), in particular the abrupt decrease in D. lodoensis from common at 316 ft (96.32 m) to absent at 321 ft (97.84 m), and preservation is suggestive of a stratigraphic gap between Zones NP12 and NP11.
The interval between 361 and 446 ft (110.03 and 135.94 m) is difficult to interpret. In contrast, two unconformities are easily inferred between 466 and 471 ft (142.04 and 143.56 m; Zone NP6 and Subzone NP4b) and between 494.7 and 501.0 ft (150.78 and 152.70 m; Zone NP2). It is likely that the Zone NP8/NP7/NP6 boundaries also represent unconformable contacts.
Cretaceous
We obtained 100 samples from the Sea Girt corehole for Cretaceous nannofossil studies, which compose an average ~10 ft (3 m) sampling interval for the 1090.5 ft (332.38 m) of Cretaceous section (from 509.5 ft to TD [155.30 m to TD]). No samples were obtained for nannofossil study, however, from the nonmarine Magothy or Potomac Formations and a closer spaced sampling was done for the Navesink and Merchantville Formations. Smear slides were prepared at Pennsylvania State University in the laboratory of T. Bralower using standard techniques and examined by L. deRomero under the light microscope at x1250 magnification.
The nannofossil zonation and CC terminology of Sissingh (1977) were used to divide the section. Several of the Sissingh (1977) zones have been modified by Perch-Nielsen (1985), based largely on reclassification of the names of original species and genera. Perch-Nielsen (1985) also defined several zonal subunits, and these are also applied here. Zones defined by Bralower (1988) have been applied to the Cenomanian/Turonian boundary interval. All zones are correlated to stages according to the scheme of Gradstein et al. (1995).
Santonian–Maastrichtian calcareous nannofossils are generally abundant and common, allowing biostratigraphic zonation. Samples from 510 to 525 ft (155.45 to 160.02 m) contained Nephrolithus frequens and were assigned to upper Maastrichtian Zone CC26. Samples between 530 and 555 ft (161.54 and 169.16 m) were assigned to middle Maastrichtian Zone CC25. The sample at 560 ft (170.69 m) contained Reinhardtites levis and Quadrum trifidum and is assigned to Subzone CC23a. The highest occurrence of Broinsonia parca at 565 ft (172.21 m) indicates the top of Subzone CC23a. The absence of Zone CC24 could indicate an unconformity between 555 and 560 ft (169.165 and 170.69 m); however, both of the Zone CC23 samples (upper Campanian) are from the basal Maastrichtian Navesink Formation and probably reflect reworking from the underlying Mount Laurel Formation. The sample at 570.4 ft (173.86 m) is assigned to Subzone CC22b/c, based on the highest occurrence of Reinhardtites anthophorus. The sample at 581 ft (177.09 m) is assigned to upper Campanian Subzone CC22a based on the HO of Quadrum grillii and the LO of Q. trifidum. Samples between 591 and 621.3 ft (180.14 and 189.37 m) are assigned to middle Campanian Zone CC21. The LO of Ceratolithoides aculeus at 721 ft (219.76 m) indicates that samples from 631 to 721 ft (192.33 to 219.76 m) are assigned to middle Campanian Zone CC20, based on the presence of C. aculeus. Samples from 730.3 to 891 ft (222.60 to 271.58 m) are assigned to lower Campanian Zone CC19 based on the HO of Marthasterites furcatus at 901.0 ft (274.62 M). Samples from 901 to 961 ft (274.62 to 292.91 m) are assigned to Zone CC18, from the HO of M. furcatus to the LO of Aspidolithus parca. Samples from 964.4 to 1060 ft (293.95 to 323.09 m) were initially assigned to upper Santonian Zone CC17 based on the absence of Eprolithus florilus. Because of the rare occurrences of holococcoliths in the interval from 1030 to 1065 ft (313.94 to 324.61 m) at Sea Girt, zonal assignment is less certain. Based on reevaluation and comparison with Bass River, samples from 1001.0 to 1050.6 ft (305.10 to 322.05 m) are assigned to Zones CC16–CC17 based on the presence of Lucianorhabdus cayeuxi and the absence of Lithastrinus moratus, which is present in Zone CC15 in Bass River. The presence of Ahmuellerella regularis throughout the interval indicates a probable assignment to upper Zone CC16 or higher. Samples from 1060 and ~1291 ft (323.09 and 393.50 m) were not examined for calcareous nannofossils.
Nannofossil biostratigraphy places constraints on the Cenomanian–Turonian Bass River Formation. Samples from the upper Bass River Formation are barren (1290.7–1356.0 ft; 393.41–413.31 m). The section from 1356 to 1417.5 ft (413.31 to 432.05 m) is assigned to uppermost Cenomanian–lower Turonian Subzone CC10b to Zone CC11 based on the presence of Gartnerago segmentatum and Isocrystallithus compactus and the absence of Lucianorhabdus maleformis and Eiffellithus eximius. The Cenomanian/Turonian boundary is tentatively placed at 1417.5 ft (432.05 m), between samples at 1415 and 1420 ft (431.25 and 432.82 m), based on the HO of Microstaurus chiastius and Lithraphidites acutus. Sample 1420.0 ft (432.82 m), however, is the only one in which M. chiastius and L. acutus were found and also has the best preservation. Thus, the Cenomanian could extend above 1417.5 ft (432.05 m). The section from 1417.5 to 1446.5 ft (432.05 to 440.89 m) is assigned to upper Cenomanian Subzone CC10a based on the HO of M. chiastius and L. acutus. Samples from 1446.5 to 1506.2 ft (440.89 to 459.09 m) are barren.
A total of 19 samples were analyzed from the Sea Girt corehole for palynomorphs, pollen, and spores (Table T12) ranging in age from Neogene to lower Cenomanian. Specimens in samples analyzed above 1106.7 ft (337.32 m) were generally poorly preserved and were not useful for biostratigraphic zonation. Samples below 1106.7 ft (337.32 m; Santonian and older) had better preservation and generally contained biostratigraphically useful pollen. Samples are assigned ages using the zonations of Brenner (1967) and Doyle and Robbins (1977).
Four samples were analyzed from the Cape May and Kirkwood Formations (30, 64.5, 103, and 120 ft; 9.14, 19.66, 31.39, and 36.58 m). Preservation in these samples was generally poor. The samples contained Miocene–Holocene pollen.
We analyzed 15 samples to help determine the age relationships of the Magothy, Bass River, and Potomac Formations. Samples at 1070.6 and 1080.5 ft (326.32 and 329.34 m) contained only fusain and did not contain palynomorphs. Samples at 1106.7 and 1136.3 ft (337.32 and 346.34 m) are assigned to Zone VII based on the presence of Heidelbergipollis sp. A and Trudopollis sp. B. Samples from 1183.2 and 1241.2 ft (360.34 and 378.32 m) contain Poropollenites sp., Complexipollis sp. E, and Complexipollis sp. B and are assigned to Zone V (equivalent to the South Amboy Fire Clay in outcrop). Samples at 1263.6 and 1284.5 ft (385.15 and 391.52 m) were barren. Samples from 1317.0 to 1410.7 ft (401.42 to 429.98 m) are assigned to pollen Zone IV. The sample at 1317.0 ft (401.42 m) contains Atlantopollis sp., typical of the Woodbridge Clay Member of the Raritan Formation (the updip equivalent of the Bass River Formation). The sample at 1410.7 ft (429.98 m), although possibly brackish water, contains Complexipollis sp. P, an index form for the Raritan Formation in outcrop. The boundary between Zones IV and III is uncertain because the sample at 1511 ft (460.55 m) is almost barren. The sample at 1541.3 ft (469.79 m) contains a well-preserved flora assigned to Zone III, based on the occurrence of Tricolpites vulgaris and Tricoporites subtlus. The lowest sample, examined at 1560.3 ft (475.58 m), contains no diagnostic pollen.
