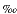 for the
for the  18O.
18O.
The sediments at Site 1079 consist of uniform olive-gray silty clays with varying amounts of nannofossils and foraminifers. In contrast, diatoms, silicoflagellates, and radiolarians are absent. The content of organic matter averages 3 wt%, which is high for ocean margin areas. We examined samples from Hole 1079A. The sampling spacing was 20 cm from 0 to 16.45 mbsf and 40 cm from 16.45 to 47.05 mbsf. Since Deep Sea Drilling Project (DSDP) Leg 94 (Ruddiman, et al., 1987), researchers have reported coring gaps between successive advanced hydraulic piston corer cores, ranging from 0.5 to 2.7 m (Ruddiman, Sarnthein, Baldauf, et al., 1988; Farrell and Janecek, 1991; Hagelberg et al., 1995). Hence, it is reasonable to consider that there might be gaps between cores. To obtain a better estimate for depth below seafloor for each sample, we assumed a gap size of 0.6 m and assigned a new depth to each sample. The coring gap was deduced by visual comparison of magnetic susceptibility and digital color reflectance data for Holes 1079A, 1079B, and 1079C (Shipboard Scientific Party, 1998b). Approximately 15 cm3 of each sample (N = 150) was freeze-dried, soaked in a Calgon solution overnight, and then washed over a 63-µm mesh sieve. The coarse fraction was dry sieved over a >150-µm mesh sieve, and in most cases, samples were split using an Otto microsplitter. Between 70 and 2000 (mean = 380) benthic foraminifers were counted from the >150-µm fraction. The >150-µm size fraction has been used extensively for paleoceanographic studies and for many studies of living foraminiferal distributions in the South Atlantic (e.g., Mackensen et al., 1995; Schmiedl, 1995). Thus, it is convenient to use the >150-µm size fraction to make direct comparisons between studies. However, examining only the larger size fraction may result in a significant loss of information, especially in intense upwelling areas and low-oxygen environments (e.g., Sen Gupta et al., 1987; Sen Gupta and Machain-Castillo, 1993). According to growth experiments, foraminiferal species mature faster with more available food (e.g., Bradshaw, 1961), but it is not clear that adult test sizes are always smaller as a result (Pedersen et al., 1988). Also, specimens in the 63- to 150-µm size fraction are more susceptible to postdepositional destruction (e.g., Rathburn and Miao, 1995).
To make our data set comparable to previous studies on living benthic foraminifers in the South Atlantic Ocean, we followed the taxonomic concepts of Mackensen et al. (1990, 1993) and Schmiedl (1995). Planktonic foraminifers and planktonic foraminiferal fragments were also counted in the same set of samples. In several samples where benthic foraminifers abundances were either very high or low, planktonic foraminifers were also identified. The relative abundance of Globigerina bulloides was used as an additional tool to recognize variations in productivity.
Organic carbon (Corg) was determined as the difference between total carbon concentrations measured by a Perkin Elmer 2400 CHN elemental analyzer (Verardo et al., 1990) and the carbonate carbon concentrations measured by a Coulometrics inorganic carbon analyzer (Engleman et al., 1985). Repeated measurements of various samples indicate that the analytical precision is ±0.87% for total carbon and ±3.3% for CaCO3.
Isotopic determinations were carried out at the Stable Isotope Laboratory of Scripps Institution of Oceanography using a Carousel-48 automatic carbonate preparation device coupled to a Finnigan MAT252 mass spectrometer. The long-term precision of the standard NBS-19 over the period of a year is better than 0.1 for the 18O.
The age model (Fig. F2A) was derived from the 18O record of Globobulimina spp., using the age assignments of Imbrie et al. (1984). These assignments are employed here for the sake of comparison with other late Quaternary records tied to the SPECMAP scale, although they are too young for both Termination I and Termination II (by 2-6 k.y.) (Berger et al., 1996). The benthic and planktonic foraminiferal accumulation rates (BFAR and PFAR) were calculated from the product of the sedimentation rates (in centimeters per thousand years), the dry bulk density (in grams per cubic centimeter), and the number of benthic and planktonic foraminifers per gram of dry sediment (in number of benthic foraminifers per gram [BF/g] and number of planktonic foraminifers per gram [PF/g]), respectively. In this study, we used BFAR as a paleoproductivity proxy (Herguera and Berger, 1991; Herguera, 1992). This method is based on the hypothesis that the number of benthic foraminifers per unit area and unit time depends on the supply of organic carbon to the seafloor. Sedimentation rates, ranging from 5 to 50 cm/k.y., were determined by linear interpolation between age control points (Fig. F2A). Dry bulk density values were interpolated from shipboard physical properties data for Site 1079 (Wefer, Berger, Richter, et al., 1998). The benthic and planktonic foraminiferal abundances vary over large ranges; some values are very small, whereas some are very large. We transformed the scale to logarithms to base 10 to enable all the data to be properly viewed.
Patterns of species association were explored through cluster analysis of the relative abundance of species with >5% in at least five samples. Clusters were based on the correlation matrix (distance metric is 1-Pearson correlation coefficient), using average-linkage clustering (SYSTAT). Spectral analyses of the number of benthic foraminifers per gram and BFAR records were performed using the software program described in Schulz and Stattegger (1997).
![]()