 18O values for pure calcite and aragonite precipitated in isotopic oxygen equilibrium with bryozoan's ambient seawater, using the following method.
18O values for pure calcite and aragonite precipitated in isotopic oxygen equilibrium with bryozoan's ambient seawater, using the following method.
Isotopic composition of living and dead bryozoan skeletons has been measured by several studies. Bone and James (1997) noted the isotopic composition of Adeonellopsis and Nevianipora from the Lacepede Shelf, east of our study area. We evaluate reliability of the bryozoan isotopes as a proxy for their inhabiting environment by examining their data. Bone and James (1997) reported summer and winter temperature profiles of water column and isotopic composition of surface water on the Lacepede Shelf but did not give any data on salinity. According to oceanographic data from LEVITUS94 (Levitus et al., 1994; Levitus and Boyer, 1994), spatial differences in mean annual salinity and mean annual temperatures of surface water are negligibly small (~0.15° and 0.4°C, respectively) at the sites where Bone and James (1997) collected Adeonellopsis and Nevianipora. Salinity and temperatures decrease uniformly with depth at those sites.
We calculated the equilibrium field of 18O values for pure calcite and aragonite precipitated in isotopic oxygen equilibrium with bryozoan's ambient seawater, using the following method.
18O of surface water is presumed to be 0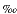 VSMOW.18O values of bottom water are estimated using the salinity profiles and relationships between changes in 18O of water and those in salinity (
VSMOW.18O values of bottom water are estimated using the salinity profiles and relationships between changes in 18O of water and those in salinity (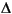 18OW/ = 0.50) (Broecker, 1989).
18OW/ = 0.50) (Broecker, 1989).
Then, 18O of equilibrium pure calcite was estimated by an equation of Friedman and O'Neil (1977) using temperature data from LEVITUS94. All of our isotopic values were corrected (18Ocorr and 13Ccorr) for the following fractionations:
Aragonite is enriched in 18O by 0.6 compared to calcite precipitated under the same conditions (Tarutani et al., 1969). The 18O in magnesian calcite is higher relative to pure calcite by 0.06 for each mol% MgCO3 in the calcite (Tarutani et al., 1969). Aragonite is enriched in 13C by 1.7 compared to coprecipitated calcite (Romanek et al., 1992).
It is clearly shown that Adeonellopsis and Nevianipora precipitate their skeleton nearly in oxygen isotopic equilibrium with ambient seawater (Fig. F7). Consequently, it is plainly evident that 18O of these bryozoans is a useful proxy as oceanographic conditions where the bryozoans grew.
Carbon isotopic composition of the equilibrium calcite and aragonite is not calculated because little is known on 13C of dissolved inorganic carbon of bottom water in the Lacepede Shelf.
If diagenesis has occurred, then the isotopic composition of the bryozoans from Holes 1129C, 1131A, and 1132B will have been changed as a result of this diagenetic alteration. A profile of chemical properties, such as Sr concentrations in interstitial water from Site 1129, indicates that the properties are rather homogenous and similar to those of ambient seawater in the interval of 0-40.3 mbsf. Consequently, this interval is thought of as a flushed zone. Sediments within this zone contain a large volume of HMC and aragonite but no dolomite (Feary, Hine, Malone, et al., 2000). Below this zone, HMC and aragonite are less common, and instead, LMC is more abundant. Bryozoans collected from 0.40 mbsf are thought to retain their original isotopic composition because their 18O and 13C values are close to those of pure calcite that is precipitated in isotopic equilibrium with ambient seawater with
18OH2O = -0.2
(calculated from salinity from LEVITUS94 data and 18OH2O values of surface water [Bone and James, 1997]) at 13.4° (mean annual temperature of bottom water [LEVITUS94 data]). Thus, bryozoan skeletons in the flushed zone may retain their original isotopic composition. Below the flushed zone, the sediments have been variably subjected to diagenesis. This conclusion is supported by two lines of evidence:
Few diagenetic signatures were observed in the uppermost 10 m of sediments in Hole 1131A, below which the sediments have been diagenetically altered to varying degrees. The flushed zone is also recognized in Hole 1132B, where the sediments in the interval 0-30.3 mbsf have been hardly subjected to diagenesis.
In this study, we investigated relationships between the mineralogy and isotopic composition of Adeonellopsis. The mineralogy is highly variable from specimen to specimen. Some skeletons consist exclusively of aragonite, and others are associated with IMC (6.6-12.1 mol% MgCO3). There is no regular relationship between mineralogy (calcite content) and 18O (Fig. F8A). It is worthwhile to note that the 18O values range widely even if the skeletal mineralogy is similar. In the case of the skeletons consisting of pure aragonite, the 18O values range from 1.43 to 2.79, with a standard deviation (1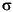 ) of 0.27 (mean = 2.17; N = 89). In contrast, the 13C values decrease with increasing calcite content (Fig. F8B). These data indicate that the original aragonitic skeletons may be accompanied by calcites that have smaller 13C and similar 18O values compared with the aragonite. Possible explanations include secondary thickening and/or diagenetic alterations.
) of 0.27 (mean = 2.17; N = 89). In contrast, the 13C values decrease with increasing calcite content (Fig. F8B). These data indicate that the original aragonitic skeletons may be accompanied by calcites that have smaller 13C and similar 18O values compared with the aragonite. Possible explanations include secondary thickening and/or diagenetic alterations.
Bryozoan oxygen isotopic profiles in Holes 1129C, 1131A, and 1132B are compared with the standard Quaternary oxygen isotopic record (Berger et al., 1994) (Fig. F9). Oxygen isotopic profiles of the three genera are generally similar to each other with some outliers. We referred to accelerator mass spectrometer (AMS) 14C and U-series ages (N.P. James et al., unpubl. data) to match the curves. The oxygen isotope profiles of Holes 1129C and 1132B can be correlated with marine isotope Stages 1-8 and those of Hole 1131A to Stages 1-4(?). Paucity of bryozoans below ~90 mbsf does not allow precise correlation with stages older than isotope Stage 9 in Holes 1129C and 1132B. It also seems that Stages 3 and 5 are not clearly discriminated in the isotopic profiles from Hole 1129C nor is Stage 2 in Hole 1132B.
Amplitudes of the profiles from Hole 1129C are smaller (<1) than those of the standard Quaternary oxygen isotopic record (<1.5), except for the large shifts at ~0-10 and 40 mbsf. Possible explanations for these include the following:
Our observations indicate that the sediments below 40 mbsf have been variably subjected to diagenesis. Thus, the latter explanation (2) is acceptable. Even though diagenesis has caused significant shifts of original bryozoan isotopic profiles and reduction of the amplitude, the shape of the profiles may be preserved, although this suggestion cannot be substantiated
Overall the bryozoan mounds correspond well to the intervals that yield bryozoans with heavier oxygen isotopic values. These intervals are correlated with marine isotope Stages 2, 4(?), 6, and 8 in Hole 1129C, Stages 2 and 4(?) in Hole 1131A, and Stages 2, 4, 6, and 8 in Hole 1132B. In contrast, bryozoan mounds did not flourish during interglacial stages, especially isotope Stages 1 and 5, as observed in Holes 1129C and 1132B. James et al. (2000) proposed a model for the mound formation at Site 1131 during the last glacial maximum. The mounds were formed by intensified upwelling and increased nutrient levels and trophic resources during glacial periods when the Leeuwin Current, which carries warm nutrient-depleted low-salinity water that forms a zone ~50 km wide and 200 m deep from off western Australia around Cape Leeuwin into the Great Australian Bight, was inactive (e.g., Wells and Wells, 1994; Wells and Okada, 1996; McGowran et al., 1997; Okada and Wells, 1997; James et al., 1999). Our data provide supporting evidence for this model.
Although Adeonellopsis spp., Idmidronea spp., and Nevianipora sp. have similar 18Ocorr values, the 13Ccorr values are quite different from one another. Nevianipora sp. has slightly lighter 13C values than Idmidronea spp. On the other hand, Adeonellopsis spp. has slightly heavier 13C values than Idmidronea spp. in Holes 1129C and 1131A, but no regular relationship is found in Hole 1132B. Figures F2, F3, and F4 show that carbon isotope fluctuation may not be related directly to glacial-interglacial oceanographic changes. Crowley and Taylor (2000) studied modern bryozoans collected from Otago Shelf, New Zealand, and showed that differences between the carbon isotopic composition of bryozoan skeletons from the same species and coexisting species living in the same community may be assignable to significant variations in the extent to which marine dissolved inorganic carbon (DIC) and respiratory CO2 are utilized during calcification. Consequently, it seems very difficult to interpret the 13C signals in Holes 1129C, 1131A, and 1132B at this time.
![]()