AGE MODEL AND SEDIMENTATION RATES
An age-depth model for the Neogene section of Hole 1138A (Fig. F6) is constructed from tephra 40Ar-39Ar ages (Table T4), biostratigraphic datums, and magnetostratigraphic tie points to the GPTS (Tables T5, T6; Fig. F6). The biostratigraphic age constraints are derived primarily from 36 diatom datums, but several planktonic foraminifer and nannofossil datums (Coffin, Frey, Wallace, et al., 2000) are also incorporated. The calibrated age for each diatom datum is compiled from the sources listed in the Table T5 caption. The age model for the Neogene section of Hole 1138A is interpreted in terms of possible error in the calibrated ages for the biostratigraphic datums (Fig. F6), and we apply a uniform ±300-k.y. error to all individual diatom (D), nannofossil (N), and planktonic foraminifer (P) datums. The line on Figure F6 represents a "best fit" age model, and the gray "envelope" represents an interpreted total uncertainty in the age model. The vertical error bars for the biostratigraphic datums (Fig. F6) reflect uncertainty in the datum depth intervals resulting from sample spacing gaps.
Three diatom zonal datums were not used in the construction of the age-depth plot for Hole 1138A. The FO of T. oliverana does not have a well-established age calibration, and there is also some taxonomic uncertainty associated with the identification of this taxon resulting from the presence of T. oliverana var. sparsa, which has a documented older/lower FO than T. oliverana s.s. (Harwood and Maruyama, 1992). Thalassiosira insigna is very rare in the upper part of its range, limiting its LO datum as a useful marker. Intermediate forms between Fragilariopsis barronii and Fragilariopsis kerguelensis are present in the upper part of the range of Fragilariopsis barronii (see Gersonde and Bárcena, 1998), which similarly inhibits accurate identification of the LO datum for F. barronii.
Four biostratigraphic datums fall outside the interpreted age envelope for Hole 1138A (Fig. F6). These include the FO of Neogloboquadrina pachyderma (P1), the LO of Cyclicargolithus floridanus floridanus (N2), the FO of Globorotalia miozea (P2), and the FO of Paragloborotalia incognita (P3). It is unclear why these datums are not consistent with the other age data. These inconsistencies may be a result of poorly calibrated ages in the southern high latitudes for nannofossils and planktonic foraminifers, sample spacing gaps, or problems with the diatom age calibrations. The coincidence of the LO of C. floridanus floridanus (N2) with diatom datums D28 and D29 (between ~191 and 186 mbsf) may indicate the presence of a middle Miocene hiatus. Well-constrained tephra ages A4 and A5 at ~171 mbsf, however, suggest the calibrated age for N2 is too young and that the middle Miocene interval is continuous.
Shipboard paleomagnetic analyses using a pass-through cryogenic magnetometer were performed on the entire sedimentary section recovered in Hole 1138A; stable and reliable inclination data were obtained after alternating-field demagnetization at 20 mT (Coffin, Frey, Wallace, et al., 2000). Although paleomagnetic data are available through the entire Neogene section, polarity reversal ties to the GPTS are only attempted in the Pliocene-Pleistocene interval. Poor recovery in the Miocene section significantly hinders paleomagnetic interpretation, particularly in intervals with many chrons of short duration (e.g., in the middle Miocene). In the Pliocene-Pleistocene section, diatom biostratigraphy provides sufficient time constraint for the recognition of Subchron C1r.1n, Chron C2n, and Subchrons C2An.1n, C2An.2n, and C3n.1n (Fig. F6; Table T6). Application of this preferred fit to the GPTS does not result in any significant inconsistencies with the biostratigraphic information. However, one exception is the placement of the top of Subchron C2An.1n. Assigning the normally polarized interval between 73.87 and 74.83 mbsf to C2An.1n requires an age assignment for the upper level (2.58 Ma at 74.83 mbsf) that is apparently too young. The reason for this inconsistency is unclear, as a hiatus does not appear to be present at this level.
Overall, a well-constrained Neogene age-depth model is derived from data compiled here for Hole 1138A. Additionally, there is excellent agreement between the biostratigraphic and radiometric ages. A relatively expanded Neogene section is present in Hole 1138A. Sedimentation rates for the Miocene section (~265-105 mbsf) averaged ~10 m/m.y., and sedimentation rates in the lower Pliocene-upper Pleistocene section (~105-0.0 mbsf) averaged ~22 m/m.y.
Compiled age information for Hole 1138A identifies an uppermost Miocene-lowermost Pliocene disconformity between Sections 183-1138A-12R-2 and 12R-3 (Fig. F6). Diatom biostratigraphy and tephra age A1 indicate this hiatus spans the interval from ~4.7 to ~5.8-6.0 Ma. The maximum age range for the hiatus is derived from the truncation of the ranges for the diatoms Nitzschia miocenica (D19) and Fragilariopsis donahuensis (D18), both with LOs between 5.8 and 6.2 Ma. This interpretation is also supported by the absence of Thalassiosira tetraoestrupii (D17) in Section 183-1138A-12R-3 (FO = 5.7 Ma). An ash age of 5.99 ± 0.71 Ma (A1) at the top of Section 183-1138A-12R-3 corroborates the biostratigraphic age interpretation. A diatom sample immediately above the ash horizon (Sample 183-1138A-12R-2, 100-101 cm; 104.80 mbsf) contains Thalassiosira complicata (D15), indicating an age of 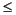 4.7 Ma. A color change is also evident in the shipboard photo of Core 183-1138A-12R. Section 183-1138A-12R-3 is slightly lighter colored than Section 12R-2 (Coffin, Frey, Wallace, et al., 2000), indicating a higher carbonate (nannofossil) content of upper Miocene sediments of Section 12R-3. Additionally, there is a magnetic polarity reversal (normal to reversed polarity in the upcore direction) between Samples 183-1138A-12R-3, 15 cm, and 12R-3, 10 cm (105.45-105.40 mbsf), further indicating the presence of a hiatus at this level.
4.7 Ma. A color change is also evident in the shipboard photo of Core 183-1138A-12R. Section 183-1138A-12R-3 is slightly lighter colored than Section 12R-2 (Coffin, Frey, Wallace, et al., 2000), indicating a higher carbonate (nannofossil) content of upper Miocene sediments of Section 12R-3. Additionally, there is a magnetic polarity reversal (normal to reversed polarity in the upcore direction) between Samples 183-1138A-12R-3, 15 cm, and 12R-3, 10 cm (105.45-105.40 mbsf), further indicating the presence of a hiatus at this level.
Another possible hiatus or condensed section is present at the Oligocene/Miocene boundary between ~270 and 260 mbsf (Fig. F6). This interval of the drill core is not well constrained biostratigraphically, due to poor core recovery and low-resolution sampling. Biostratigraphic data indicate that a hiatus, if indeed present, spans the time interval between ~22.5 and 24.0 Ma.
