BIOSTRATIGRAPHY
The Neogene section of nannofossil ooze, chalk, and clay in Holes 1209A and 1209B ranges from the Pleistocene to the lower Miocene. Hole 1209C was cored down from the middle Miocene. Two unconformities or condensed intervals are present within the upper Miocene (Samples 198-1209A-9H-CC to 10H-CC and 198-1209B-10H-CC to 11H-CC) and between the upper and lower Miocene (Samples 198-1209B-11H-CC to 12H-CC and 198-1209C-1H-CC to 2H-CC) as revealed by calcareous plankton bioevents. A second major unconformity separates the lower Miocene from the Oligocene (Samples 198-1209B-12H-CC to 13H-CC and 198-1209C-2H-CC to 3H-CC). The Eocene and the Paleocene sections were triple-cored and appear to be complete, including the P/E boundary transition, the PETM critical interval, and the K/T boundary, at least to within the calcareous plankton biostratigraphic resolution achieved shipboard. A thick and continuous middle-upper Maastrichtian sequence of white nannofossil ooze was also triple-cored at Site 1209. The main calcareous nannofossil and foraminiferal datums are summarized in Tables T2 and T3.
Calcareous nannofossils are generally abundant and moderately to well preserved throughout the Cenozoic and Mesozoic sediments of Holes 1209A, 1209B, and 1209C. Neogene planktonic foraminifers are moderately to well preserved, but foraminiferal abundance is relatively low, due to selective dissolution and fragmentation. Dissolution and generally poor preservation characterizes the Oligocene and upper and middle Eocene assemblages, whereas the preservation improves slightly in the remainder of the Eocene and Paleocene section. Cretaceous planktonic foraminifers are well preserved and abundant. All core catcher samples were examined and supplementary samples used to refine datums and the stratigraphy in and around critical intervals.
Studies on benthic foraminifers were conducted on selected core catcher samples from Holes 1209A and 1209C. The >250-µm size fraction was examined for the Neogene samples, and the >125-µm size fraction was examined for the Paleogene and Cretaceous samples. Neogene benthic foraminifers are well preserved and common; Paleogene and Cretaceous taxa are moderately well preserved but rare.
Neogene
The Pleistocene and Neogene section ranges from the upper Pleistocene (Subzone CN14b) to the lower-middle Miocene (Zones CN1 to CN3). Calcareous nannofossils are generally abundant and moderately to well preserved throughout, and most of the zones of Okada and Bukry (1980) were recognized. Zones CN8, CN6, and CN5 were not identified, most likely due to condensed intervals and/or short unconformities; calculations reveal a marked decrease in sedimentation rate around 7 Ma in Zone CN9.
The middle to lower Miocene (Zones CN3 to CN2) is difficult to subdivide, due to the sporadic occurrence of the nominal taxon Sphenolithus heteromorphus, as reported from previous Leg 198 sites. At Site 1209, S. heteromorphus was recorded from Samples 198-1209B-12H-CC and 198-1209C-2H-CC. A lower Miocene interval (Zones CN1 to CN3) was identified based on the presence of abundant Discoaster deflandrei (acme top within Zone CN3) and Cyclicargolithus floridanus, the absence of S. heteromorphus and Calcidiscus premacintyrei, and the absence of older, robust taxa such as Cyclicargolithus abisectus, Reticulofenestra bisectus, and Zygrhablithus bijugatus, which have LOs in Subzone CN1a and lower Subzone CN1b.
Paleogene
An unconformity and/or condensed interval separates the lower Miocene (Zone CN1 to CN3) and lower Oligocene (Subzone CP19a), but the lower Oligocene and Oligocene-Eocene transition appears to be complete.
The Eocene and Paleocene section also appears to be complete, including the Paleocene/Eocene boundary transition and the PETM critical interval. Events constraining this interval include the abundance decline and last occurrence of the genus Fasciculithus (e.g., interval 198-1209B-22H-1, 12-134 cm), the FO of Discoaster diastypus (e.g., Sample 198-1209B-21H-6, 120 cm; base of Subzone CP9a), and the FO of Tribrachiatus orthostylus (e.g., Sample 198-1209B-21H-4, 100 cm; base of Subzone CP9b). Other biostratigraphically significant species of the Tribrachiatus lineage are also present but are not well preserved.
The K/T boundary also appears to be complete and includes a relatively expanded series of datum events following the LO of Cretaceous taxa (see Fig. F3). The boundary is overlain by a 16-cm carbonate layer dominated by fine micrite and abundant calcispheres and tiny foraminifers, with rare to few reworked Cretaceous nannofossil taxa and frequent survivor taxa (e.g., Cyclagelosphaera and Markalius inversus) (e.g., Samples 198-1209A-25H-6, 90 and 94 cm) (see Fig. F3). This is followed by influxes of Neobiscutum (Sample 198-1209A-25H-6, 40 cm) and, subsequently, Cruciplacolithus (Sample 198-1209A-25H-6, 10 cm).
Cretaceous
An apparently continuous section of Maastrichtian sediments was recovered from Site 1209. The calcareous nannofossils from this interval are well preserved and the assemblages diverse. Zones CC26 to CC24 were identified; however, the nominal taxon Lithraphidites quadratus was rare and sporadically distributed, and Tranolithus orionatus was very rare. We had difficulty identifying the LO of Reinhardtites levis. Therefore, there is a degree of uncertainty in our datum depths, and this is borne out by the position of these datums with respect to the best-fit sedimentation rate curve (see Fig. F22).
Neogene
The Neogene section ranges from upper Pleistocene to middle Miocene, with two possible unconformities separating parts of the upper and middle Miocene; another unconformity separates the middle Miocene and lower Oligocene. Planktonic foraminifers at Site 1209 are generally moderately preserved. They are abundant to common in the Pleistocene and Pliocene but then decline in abundance and preservation downward through the Miocene.
The youngest sediments recovered in Holes 1209A and 1209B are assigned to Pleistocene Zone N22 based on the co-occurrence of Truncorotalia tosaensis and Truncorotalia truncatulinoides in Samples 198-1209A-1H-CC and 2H-CC and 198-1209B-1H-CC to 3H-CC. Common Pleistocene taxa include Globigerina bulloides, Globorotalia inflata, Globorotalia crassaformis, Orbulina universa, Neogloboquadrina dutertrei, and Neogloboquadrina pachyderma (dextral). These are supplemented by few Globorotalia menardii, Globorotaliatumida, and pulleniatinids. Within the Pliocene sediments the FOs of T. tosaensis, T. crassaformis (Samples 198-1209A-6H-CC and 198-1209B-5H-CC), and Sphaeroidinella dehiscens (Samples 198-1209A-7H-CC and 198-1209B-8H-CC) allow the differentiation of Zones N21 and N20-N19, respectively. Other useful bioevents are the LOs of Dentoglobigerina altispira (Samples 198-1209A-6H-CC and 198-1209B-8H-CC), Sphaeroidinellopsis seminulina (Samples 198-1209A-5H-CC and 198-1209B-5H-CC), and Globoturborotalita nepenthes (Samples 198-1209A-7H-CC and 198-1209B-8H-CC).
The Miocene sequence at Site 1209 is interrupted by unconformities, and planktonic foraminiferal preservation is slightly improved compared with Sites 1207 and 1208. The uppermost Miocene is present in Sample 198-1209B-8H-CC, which contains common Globorotalia tumida, Globorotalia conoidea, Globorotalia miozea, Globoturborotalita nepenthes, and Sphaeroidinellopsis seminulina. This sample is assigned to Zone N18 based on the presence of G. tumida and the absence of Sphaeroidinella dehiscens. The presence of common late Miocene taxa such as Globorotalia plesiotumida, Dentoglobigerina altispira, Globoquadrina venezuelana, G. nepenthes, and G. conoidea (from Samples 198-1209A-9H-CC to 10H-CC and 198-1209B-9H-CC) permit the identification of Zone N17 based on the absence of G. tumida. A minor unconformity separates this interval from the remainder of the upper Miocene sequence drilled in Holes 1209A (Samples 198-1209A-10H-CC and 11H-CC) and 1209B (Sample 198-1209B-11H-CC), which yield few to common Globoquadrina dehiscens, G. nepenthes, Globoturborotalita woodi, Globoturborotalita decoraperta, and Dentoglobigerina altispira. In the absence of Neogloboquadrina acostaensis and based on the presence of G. nepenthes, the assemblages are attributed to Zones N14 to N15. This interval overlies middle Miocene sediments recovered in Samples 198-1209B-12H-CC and 198-1209C-1H-CC, which contain Fohsella peripheroronda, G. dehiscens, Sphaeroidinellopsis disjuncta, Globigerinoides mitra, and Praeorbulina glomerosa and indicate Zones N8 and N9.
Paleogene
An unconformity separates the Miocene and Oligocene at Site 1209. The paucity of planktonic foraminiferal marker species renders age determination problematic for much of the Oligocene examined at this site. Dissolution is a likely factor for the absence of many taxa based on the abundance of foraminiferal fragments and crystals of phillipsite in Oligocene age samples. For example, Sample 198-1209B-13H-CC contains the robust taxa Catapsydrax dissimilis, Catapsydrax unicavus, "Globigerina" euapertura, and "Globoquadrina" pseudovenezuelana, but no zonal marker species are present. A similar assemblage is present in Sample 198-1209C-2H-CC; however, the presence of Subbotina angiporoides without Pseudohastigerina spp. indicates Zones P19 and P20. The presence of numerous specimens of Globoquadrina dehiscens in this sample suggests that downhole contamination is a problem. Lower Oligocene Zone P18 is identified in Sample 198-1209A-13H-CC based on the presence of Pseudohastigerina spp. together with Cassigerinella chipolensis, "Globigerina" ampliapertura, Tenuitella gemma, Tenuitellinata angustiumbilicata, Catapsydrax spp., "Globoquadrina" pseudovenezuelana, and Subbotina gortanii.
Dissolution and generally poor preservation continues down into the upper Eocene. Sample 198-1209A-14H-CC is assigned to Zones P16 to P17 based on the presence of "Globigerina" ampliapertura, spines of Hantkenina, and poorly preserved specimens of Hantkenina alabamensis. The sample is dominated by dissolution-resistant taxa, including Globigerinatheka senni, Catapsydrax unicavus, and "Globigerina" euapertura. A similar assemblage, including H. alabamensis, is present in Sample 198-1209B-14H-CC. Sample 198-1209C-4H-CC contains a nearly monogeneric assemblage of Globigerinatheka spp.
Preservation improves somewhat in the upper part of the middle Eocene. Sample 198-1209B-15H-CC contains an assemblage indicative of Zones P12 to P15 that includes Globigerinatheka index, Globigerinatheka mexicana, G. senni, Turborotalia pomeroli, C. unicavus, and spines of Hantkenina. Zone P13 is based on the total range of "Orbulinoides" beckmanni; rare specimens of this taxon are present in Sample 198-1209A-15H-CC together with G. index, G. mexicana, G. senni, Acarinina bullbrooki, and Acarinina primitiva. Middle Eocene Zone P11 is present in Samples 198-1209A-16H-CC, 198-1209B-16H-CC, and 198-1209C-6H-CC. This zone is characterized by the presence of Morozovella aragonensis, A. bullbrooki, Igorina broedermanni, Globigerinatheka index, Globigerinatheka subconglobata, Hantkenina mexicana, and Guembelitrioides higginsi. The co-occurrence of Hantkenina (observed only as isolated spines) together with Morozovella caucasica and Acarinina pentacamerata serves to distinguish Zone P11 from P10. The FO of Hantkenina defines the Zone P9/P10 boundary.
Lower Eocene Zones P8 and P9 are not differentiated because of the absence of Planorotalites palmerae. Samples 198-1209A-18H-CC and 19H-CC, 198-1209B-17H-CC and 18H-CC, and 198-1209C-7H-CC and 8H-CC are assigned to Zones P8 to P9. These samples are characterized by Morozovella aragonensis, M. caucasica, Acarinina pentacamerata, Acarinina soldadoensis, Igorina broedermanni, and Acarinina quetra, as well as Globigerinatheka subconglobata micra in the upper part and M. subbotinae in the lower part of this interval. Zone P7 is distinguished from the overlying sediments by the co-occurrence of M. aragonensis and Morozovella formosa and by the absence of A. pentacamerata in Samples 198-1209A-20H-CC, 198-1209B-20H-CC, and 198-1209C-9H-CC. The boundary between Zones P6 and P7 is present between Samples 198-1209A-21H-3, 68-69 cm, and 21H-4, 129-130 cm. Characteristic taxa of Zone P6 include M. subbotinae, M. aequa (in Subzone P6a), M. formosa, M. gracilis, Subbotina velascoensis, and A. soldadoensis in the absence of both M. aragonensis and M. velascoensis.
The extinction level of Morozovella velascoensis is used to define the P6/P5 zonal boundary, as well as the Paleocene/Eocene epoch boundary, and this datum occurs between Samples 198-1209A-21H-5, 49-50 cm, and 21H-5, 129-130 cm. The PETM (Zachos et al., 1993) and benthic foraminiferal extinction (BFE) event (Thomas and Shackleton, 1996) occur within Zone P5, and the interval is identified in Section 198-1209A-21H-6 based, in part, on the presence of the benthic foraminifer Gavelinella beccariiformis in Sample 198-1209A-21H-CC, a taxon that goes extinct at the BFE. Zone P5 contains a rich assemblage of M. velascoensis, Morozovella occlusa, M. subbotinae, M. aequa, Morozovella pasionensis, and Acarinina soldadoensis. The total range of Globanomalina pseudomenardii defines upper Paleocene Zone P4, which co-occurs with a diverse assemblage, including M. aequa (in the upper part), M. velascoensis, M. occlusa, M. pasionensis, Igorina pusilla, Igorina albeari, Igorina tadjikistanensis, Acarinina mckannai, Acarinina nitida, and Acarinina subsphaerica (in the lower part). Samples 198-1209A-22H-CC and 23H-CC, 198-1209B-22H-CC, and 198-1209C-11H-CC are assigned to Zone P4. Morozovella angulata, M. conicotruncata, Subbotina triloculinoides, and Parasubbotina pseudobulloides mark Zone P3 in Sample 198-1209C-13H-CC.
Lower Paleocene Zone P2 (Samples 198-1209A-24H-CC, 198-1209B-24H-CC, and 198-1209C-14H-CC) contains Praemurica uncinata, Praemurica inconstans, Morozovella praeangulata, and Parasubbotina pseudobulloides. The interval containing Zone P1 and its subzones is thin at Site 1209 and was not observed in core catcher samples. However, several samples taken across the K/T boundary permit the recognition of the Parvularugoglobigerina eugubina Zone (Zone P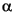 ) in the interval between Samples 198-1209A-25H-6, 93-94 cm, and 113-114 cm, and between Samples 198-1209C-15H-3, 67-68 cm, and 85-86 cm (see Fig. F3). The assemblage includes the species P. eugubina, Parvularugoglobigerina extensa, woodringinids, and Parasubbotina pseudoinconstans.
) in the interval between Samples 198-1209A-25H-6, 93-94 cm, and 113-114 cm, and between Samples 198-1209C-15H-3, 67-68 cm, and 85-86 cm (see Fig. F3). The assemblage includes the species P. eugubina, Parvularugoglobigerina extensa, woodringinids, and Parasubbotina pseudoinconstans.
Cretaceous
Cretaceous planktonic foraminifers recovered at Site 1209 are generally abundant and well preserved. The highest Cretaceous assemblages are present in Samples 198-1209A-25H-6, 113-114 cm, 198-1209B-26H-CC, and 198-1209C-15H-3, with few Paleocene taxa indicative of the P0 and P Zones in the burrows (see Fig. F3). The large-sized Cretaceous taxa include Abathomphalus mayaroensis, Pseudoguembelina costulata, P. excolata, and P. hariaensis, as well as the small taxa Schackoina cenomana, Globigerinelloides subcarinatus, Globotruncanella havanensis, and Globotruncanella petaloidea, indicating the upper Maastrichtian A. mayaroensis Zone (KS31).
The underlying Upper Cretaceous interval (Samples 198-1209A-25H-6, 123-124 cm, to 28H- CC, 198-1209B-27H-CC to 28H-CC, and 198-1209C-16H-CC to 18H-CC) is characterized by a moderately to well-preserved planktonic foraminiferal assemblage yielding Abathomphalus mayaroensis, Abathomphalus intermedius, Contusotruncana contusa, Racemiguembelina fructicosa, Pseudotextularia elegans, common pseudoguembelinids, Globotruncanita stuarti, Globotruncanita stuartiformis, Globotruncana orientalis, Heterohelix rajagopalani, and rugoglobigerinids. Based on the presence of A. mayaroensis this interval is still assigned to the upper Maastrichtian A. mayaroensis Zone (KS31).
The older Cretaceous sequence has been recovered only in Holes 1209B and 1209C. Samples 198-1209B-29H-CC to 30H-CC and from 198-1209C-19H-CC to 20H-CC yield common Contusotruncana contusa and Racemiguembelina fructicosa in the absence of A. mayaroensis, indicating that this interval belongs to the lower Maastrichtian C. contusa-R. fructicosa Zone (upper KS30). Other species present in this interval are Contusotruncana patelliformis, Globotruncana bulloides, Globotruncana linneiana, G. stuarti, G. stuartiformis, Rugotruncana subcircumnodifer associated with rare Gansserina gansseri and Gansserina wiedenmayeri, and common pseudoguembelinids and rugoglobigerinids. The lowest parts of Holes 1209B (Samples 198-1209B-31H-CC to 32H-CC) and 1209C (Samples 198-1209C-21H-CC to 22H-CC) are assigned to the lower Maastrichtian-upper Campanian G. gansseri Zone (KS30), based on the absence of C. contusa and R. fructicosa and the presence of few G. gansseri.
Benthic foraminifers were examined in selected core catcher samples from Holes 1209A and 1209C. In these holes, Neogene sediments contain generally well-preserved and rare to few benthic foraminifers. In Samples 198-1209A-25H-CC and 198-1209C-3H-CC and 25H-CC, planktonic foraminifers are strongly dissolved, and consequently, the relative abundance of benthic foraminifers is higher. The test size of benthic foraminifers is generally small in the Paleogene- and Cretaceous-age sediments, so the >125-µm size fraction was examined in these intervals. Distributions of the Cenozoic and Cretaceous benthic foraminifers are plotted in Tables T4 and T5.
Neogene
Samples 198-1209A-1H-CC to 2H-CC yield Oridorsalis umbonatus, Pullenia bulloides, uvigerinids, and Pyrgo murrhina. In Samples from 198-1209A-5H-CC to 9H-CC, Oridorsalis tener, Pullenia bulloides, Stilostomella subspinosa, uvigerinids, gyroidinoidids, and cibicidoidids characterize the benthic assemblage, except for Sample 198-1209A-7H-CC, in which uvigerinids are absent. The stilostomellids become more frequent in Sample 198-1209A-11H-CC.
Sample 198-1209C-1H-CC, which is Miocene in age, contains common calcareous trochospiral forms such as Anomalinoides semicribratus, Gyroidinoides girardanus, Oridorsalis tener, and Pullenia bulloides and the agglutinated taxa Martinottiella sp. and Eggerella bradyi. Uvigerinids and Pyrgo murrhina are present as a minor component of the assemblage in this sample.
Paleogene
Bulimina trinitatensis, Oridorsalis umbonatus, nodosariids, stilostomellids, Vulvulina mexicana, and V. spinosa are the main components in Sample 198-1209A-12H-CC and 13H-CC. Uvigerinids that characterized the Neogene section are absent.
The preservation of benthic foraminifers is poor to moderate in the lower Oligocene to lower Eocene (Samples 198-1209A-14H-CC to 20H-CC), and their test size is generally small. Buliminids (Bulimina impendens, Bulimina jarvisi, Bulimina semicostata, B. trinitatensis, and Quadratobuliminella pyramidata) are the characteristic group in this interval. Stilostomellids are also present, but they decline in abundance. The 250-µm size fraction of Sample 198-1209A-21H-CC, which is late Paleocene in age, is characterized by the dominance of agglutinated forms such as Tritaxia spp., Pseudoclavulina rugolosa, and Spiroplectammina jarvisi, whereas Gavelinella beccariiformis and Conorbinoides hillebrandti are common in the 125- to 250-µm size fraction.
Samples 198-1209A-23H-CC and 24H-CC are dominated by Nuttallides truempyi, Oridorsalis umbonatus, Aragonia velascoensis, and buliminids (Bulimina trinitatensis, Bulimina velascoensis, and Buliminella grata). Marssonella trochoides commonly occurs in Sample 198-1209A-24H-CC. In Samples 198-1209C-3H-CC and 5H-CC, buliminids (Bulimina impendens, B. jarvisi, and B. semicostata) and the stilostomellids (Stilostomella spp., Stilostomella abyssorum, Stilostomella gracillima, and S. subspinosa) characterize the benthic assemblage. The virtual absence of the calcareous trochospiral morphotypes in Sample 198-1209C-5H-CC is notable. In Samples 198-1209C-7H-CC through 10H-CC, trochospiral forms (N. truempyi, O. umbonatus, and O. tener), buliminids such as Bulimina glomarchallengeri, B. jarvisi, and B. semicostata and species of Stilostomella represent the main components of the benthic assemblage. Chrysalogonium breviloculum is common in Sample 198-1209C-9H-CC. Oridorsalis umbonatus and N. truempyi are the main components in the interval from Sample 198-1209C-11H-CC to 13H-CC. Agglutinated taxa (M. trochoides, Spiroplectammina jarvisi, and Pseudoclavulina rugolosa) rarely occur in this interval.
Cretaceous
Maastrichtian Samples 198-1209A-25H-CC and 27H-CC contain common Nuttallides truempyi and Oridorsalis umbonatus as well as Aragonia velascoensis, Lenticulina spp., and Nodosaria spp. Pseudoclavulina rugolosa is common in the >250-µm size fraction of Sample 198-1209A-25H-CC.
In Samples 198-1209C-15H-CC to 25H-CC (Maastrichtian), the benthic assemblage is generally characterized by N. truempyi, O. umbonatus, and a variety of agglutinated species (Gaudryina pyramidata, Gaudryina spp., Marssonella trochoides, Pseudoclavulina rugolosa, Spiroplectammina jarvisi, and Tritaxia spp.), Lenticulina spp., and Dentalina spp. Other benthic foraminifers sporadically present in this interval are Conorbinoides hillebrandti, Gyroidinoides spp., Cibicidoides velascoensis, Quadrimorphina allomorphinoides, Sliterella lobulata, Gavelinella beccariiformis, and Aragonia velascoensis.
Paleodepth estimates are based on the work of Pflum et al. (1976), Tjalsma and Lohmann (1983), Woodruff (1985), and van Morkhoven et al. (1986) for the Cenozoic section. For the Cretaceous section, estimates are mainly based on the study of Nyong and Olsson (1984) and the backtracked paleodepth curve from Deep Sea Drilling Program (DSDP) and Ocean Drilling Program (ODP) data (Kaiho, 1999).
The miliolids Pyrgo murrhina and Pyrgo lucernula, and the uvigerinids Uvigerina hispida and Uvigerina peregina indicate upper abyssal (2000-3000 m) depth during the Neogene (Pflum et al., 1976; Woodruff, 1985). These taxa are associated with calcareous trochospiral species (Oridorsalis tener, Pullenia bulloides, Cibicidoides subhaidingeri, Cibicidoides wuellerstorfi, and Gyroidinoides girardanus) that exhibit a wider bathymetric range from the upper bathyal to abyssal zone (Pflum et al., 1976). In the Paleogene, benthic assemblages are generally represented by Nuttallides truempyi, Oridorsalis umbonatus, Aragonia velascoensis, and buliminids (Tjalsma and Lohmann, 1983) that also indicate deep water. The genus Aragonia typically characterizes Late Cretaceous benthic assemblages. A. velascoensis and Aragonia ouezzanensis indicate lower bathyal and abyssal paleodepths, respectively (Nyong and Olsson, 1984). In addition, Pyramidina szajnochae, a characteristic Late Cretaceous form from Section 198-1209C-22H-CC, suggests an uppermost abyssal depth (2000-2500 m) (Nyong and Olsson, 1984). Therefore, the paleodepth during the Maastrichtian is similar to that of the Paleogene. These estimates are consistent with backtracked paleodepths of Kaiho (1999).
