BIOSTRATIGRAPHY
The Neogene section of nannofossil ooze and clay-rich nannofossil ooze in Holes 1210A and 1210B ranges from the Pleistocene to lower Miocene. This interval appears to be complete, at least within the stratigraphic resolution achieved shipboard. Sedimentation rates in the Pleistocene, Pliocene, and upper Miocene are relatively high (Samples 198-1210A-1H-CC through 6H-CC and 198-1210B-1H-CC through 5H-CC) but decrease considerably in older Neogene sediments (Samples 198-1210A-6H-CC through 7H-2, 20 cm) (see "Sedimentation and Accumulation Rates"). Planktonic foraminiferal datums indicate that a significant unconformity exists near the base of the Miocene (Samples 198-1210A-13H-CC and 198-1210B-13H-CC) that extends into the lower Oligocene. However, samples taken at higher resolution for study of calcareous nannofossils reveal the possibility of a condensed section in this interval.
With the exception of the condensed/truncated mid- and upper Oligocene sediments, the Paleogene section from Site 1210 is also relatively complete. High-resolution sampling for both calcareous nannofossil and planktonic foraminiferal biostratigraphy suggests that several key intervals were recovered, including the Eocene/Oligocene boundary, the Paleocene/Eocene boundary transition, PETM, and K/T boundary. A thick and continuous section of nannofossil ooze was also recovered from the Maastrichtian to the lower part of the upper Campanian, which may contain the mid-Maastrichtian event. The main calcareous nannofossil and foraminiferal datums are summarized in Tables T3 and T4.
Calcareous nannofossils are generally abundant and moderately to well preserved in the Cenozoic and Mesozoic sediments of Holes 1210A and 1210B. Planktonic foraminifers exhibit variable preservation and diversity. In particular, preservation is generally poor in samples from the middle to lower Miocene, lower Oligocene, and upper Eocene. This is indicated by the absence of more delicate taxa, higher percentages of dissolution-resistant species, and the prevalence of authigenic minerals. All core catcher samples were examined, and supplementary samples were used to refine datums and the stratigraphy in and around critical intervals.
Studies on benthic foraminifers were conducted on selected core catcher samples from Holes 1210A and 1210B. The >250-µm size fraction was examined in Neogene samples, and the >125-µm size fraction was examined in Paleogene and Cretaceous samples. Pleistocene through Oligocene sediments yield benthic foraminifers that are well preserved and rare to few in abundance. The Eocene through Cretaceous sediments yield moderately preserved benthic foraminifers that are generally rare to few in abundance.
Calcareous nannofossils are generally abundant and moderately to well preserved in Holes 1210A and 1210B. Both the Cenozoic and Mesozoic parts of the succession appear to be stratigraphically complete, at least to within nannofossil biostratigraphic resolution achieved shipboard. However, a number of upper to middle Miocene zones are missing, and because zonal markers are extremely rare or absent, an unconformity may separate the lower or middle Miocene and lower Oligocene.
Neogene
The Neogene section ranges from the upper Pleistocene (Subzone CN14b) to the lower middle Miocene (Zones CN3-CN4). Most of the zones of Okada and Bukry (1980) were recognized. The middle to lower Miocene (Subzone CN5a to Zone CN3) was difficult to subdivide because of the sporadic presence of the nominal taxon Sphenolithus heteromorphus, as reported from previous Leg 198 sites. At Site 1210, S. heteromorphus was only recorded in Sample 198-1210B-12H-CC.
Paleogene
Between Samples 198-1210A-12H-CC and 13H-CC and 198-1210B-12H-CC and 13H-CC there is a stratigraphic gap of ~13 m.y., comprising Zone CN2 (lower Miocene) to Zone CP18 (lower Oligocene). The underlying Paleogene section appears to be relatively complete. The PETM interval is characterized by the presence of abundant Discoaster multiradiatus, the abundance decrease and last occurrence of Fasciculithus (in Section 198-1210A-20H-6), and, above, by the first occurrence of Discoaster diastypus (Sample 198-1210A-20H-3, 100 cm), which marks the base of Zone CP9.
The K/T boundary interval is apparently complete. Sediments containing Micula prinsii, the nominal taxon for Zone CC26, are overlain by a white carbonate interval, which yields smear slides dominated by fine micrite, minute foraminifers, and abundant calcispheres, with few reworked Cretaceous nannofossil taxa. These sediments consistently contain survivor taxa (e.g., species of Cyclagelosphaera and Markalius). The first new Paleocene coccolith genus, Neobiscutum, is recorded in Sample 198-1210B-24H-1, 40 cm.
Cretaceous
The apparently continuous section of Maastrichtian to Campanian sediments recovered at Site 1210 yields well-preserved and diverse assemblages of calcareous nannofossils. Zones CC26 to CC21-CC20 were identified, but a number of subzones could not be identified. Certain marker taxa were difficult to identify with certainty because of the presence of transitional morphologies (e.g., Micula murus and Reinhardtites levis) or were very rare (e.g., Tranolithus orionatus). In addition, a number of ranges that are inconsistent with other sections were recorded; Lithraphidites quadratus was not found beneath the range of M. murus, and Eiffellithus eximius did not co-occur with Uniplanarius trifidum. The nature of these difficulties will be a focus of postcruise investigations.
Neogene
Planktonic foraminifers are common to abundant, and preservation is moderate to good in the apparently complete upper Pliocene to Pleistocene interval (Zones N21-N22; Sections 198-1210A1H-CC to 6H-CC and 198-1210B-1H-CC to 6H-CC). Globorotalia inflata and/or G. crassaformis are particularly abundant through this interval.
The upper Miocene-lower Pliocene interval also appears to be complete, although abundance varies from few to common and preservation is poor to moderate. A downhole increase in foraminiferal test fragmentation indicates that dissolution may be partially responsible for the sporadic distribution of biostratigraphically useful taxa through this interval. However, the last occurrences of Dentoglobigerina altispira, Sphaeroidinellopsis seminulina, Globorotalia margaritae, and Globoturborotalita nepenthes are valuable orbitally tuned datums that are present in the mid- to lower Pliocene. Likewise, the FO of Globorotalia tumida is a reliable datum in the uppermost Miocene (base of Zone N18).
Fragmentation and very low foraminiferal abundances are prevalent through the condensed lower portion of the upper Miocene (Zones N14-N5; Samples 198-1210A-11H-CC and 198-1210B-11H-CC) and also in the middle Miocene (Zone N10; Samples 198-1210A-12H-CC and 198-1210B-12H-CC). This interval is dominated by dissolution-resistant taxa, including Sphaeroidinellopsis seminulina, S. kochii, S. disjuncta, Globoquadrina dehiscens, and G. venezuelana. Rare age-diagnostic taxa in Zone N14 include Globoturborotalita nepenthes, Paragloborotalia mayeri, Globorotalia menardii, and G. praemenardii. Rare age-diagnostic taxa in Zone N10 include Orbulina universa, Fohsella peripheroacuta, and Globigerinoides mitra.
Paleogene
Low-resolution planktonic foraminiferal evidence suggests that the middle Miocene unconformably overlies the lower Oligocene (Zone P18) at Site 1210. Alternatively, it is possible that dark-colored sediments in Cores 198-1210A-13H and 198-1210B-13H represent continuous but condensed sedimentation. Like the middle Miocene, the lower Oligocene and upper Eocene assemblages also show strong dissolution based on the abundance of foraminiferal fragments, low-diversity assemblages dominated by dissolution-resistant taxa, residues dominated by markedly smaller size fractions, and the presence of the authigenic mineral phillipsite. Samples 198-1210A-13H-CC and 198-1210B-13H-CC are assigned to lower Oligocene Zone P18 based on the presence of rare to abundant Pseudohastigerina spp. in the absence of late Eocene taxa such as Turborotalia cerroazulensis and Hantkenina spp. (including spines of this genus). Other taxa present include Cassigerinella chipolensis, "Globigerina" euapertura, Catapsydrax dissimilis, and C. unicavus, with or without rare "Globigerina" ampliapertura, T. increbescens, and Globoquadrina tapuriensis.
The upper Eocene of this site contains few planktonic foraminifers in the >63-µm size fraction. Sample 198-1210B-14H-1, 7 cm, contains rare Hantkenina spines and fragments, Subbotina angiporoides, "Globigerina" ampliapertura, and common phillipsite and calcispheres. The assemblages in Samples 198-1210A-14H-CC and 198-1210B-14H-CC contain Globigerinatheka senni, G. mexicana, and G. index and are assigned to Zones P14-P16. Tektites were found in the former sample. Samples 198-1210A-15H-CC and 198-1210B-15H-CC are assigned to middle Eocene Zone P11 based on the presence of Morozovella aragonensis, Globigerinatheka mexicana, G. subconglobata, G. senni, Acarinina bullbrooki, A. primitiva, A. pentacamerata, Guembelitrioides higginsi, and rare, isolated spines of the genus Hantkenina. Zone P10 was not recognized in the low-resolution shipboard sampling, although we believe that the entire upper and middle Eocene sequence is complete based on the combined planktonic foraminiferal and calcareous nannofossil datums depicted in the age-depth plots (see "Sedimentation and Accumulation Rates").
A more expanded lower Eocene section is present at Site 1210, a trend also observed at Site 1209. The interval from Sample 198-1210A-16H-CC through 198-1210B-17H-CC represents Zone P9, which is characterized by Morozovella aragonensis, M. caucasica, Acarinina pentacamerata, A. quetra, A. soldadoensis, and Igorina broedermanni. Zones P7 and P8 (Samples 198-1210A-18H-CC, 198-1210B-18H-CC, and 198-1210A-19H-CC) contain similar assemblages but are distinguished from the overlying sediments by the co-occurrence of M. aragonensis, M. formosa, and M. subbotinae; the FO of A. pentacamerata defines the boundary between these two zones. The section is more condensed in the basal Eocene. Sample 198-1210B-19H-CC is assigned to Zone P6 based on the absence of both M. aragonensis and M. velascoensis; the presence of M. formosa places this sample in Subzone P6b.
A continuous succession of upper Paleocene sediments was recovered in Core 198-1210A-20H through Section 23H-CC and Core 198-1210B-20H through Section 23H-CC. Zone P5 (Sample 198-1210A-20H-CC) contains a diverse planktonic assemblage including Morozovella velascoensis, M. subbotinae, M. aequa, M. occlusa, and M. pasionensis. This sample also contains the benthic foraminifer Gavelinella beccariiformis, indicating a level below the benthic foraminiferal extinction event at the onset of the PETM (Zachos et al., 1993). The absence of M. subbotinae and the presence of Globanomalina pseudomenardii, Igorina pusilla, I. albeari, Acarinina nitida, A. mckannai, and A. subsphaerica distinguish Zone P4 (Samples 198-1210A-21H-CC through 22H-CC and 198-1210B-20H-CC through 21H-CC) and its subzones. The co-occurrence of M. angulata, M. conicotruncana, and I. pusilla characterizes the sediments of Zone P3 (Samples 198-1210A-23H-CC and 198-1210B-22H-CC).
A prominent, 18 cm-thick magnetic susceptibility increase was found in Core 198-1210A-22H. This layer consists of a dark brown nannofossil ooze with clay. Preliminary micropaleontological investigations suggest that this interval as already identified in cores from Site 1209 may represent a previously unrecognized event of considerable evolutionary significance. This horizon lies within planktonic foraminiferal Zone P4 and coincides exactly with the evolutionary FO of the nannolith Heliolithus kleinpellii, an important component of the late Paleocene assemblages and a marker for the base of Zone CP5. Planktonic foraminifera are characterized by a low diversity and a largely dissolved assemblage dominated by representatives of Igorina (mainly Igorina tadjikistanensis and Igorina pusilla). The composition of the assemblage suggests some kind of oceanic perturbation.
The lower Paleocene (Danian) is condensed at Site 1210. Sediments of Zones P1 and P2 were not observed in core catcher samples. However, Parvularugoglobigerina eugubina (Zone P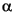 ) is present in Samples 198-1210A-24H-4, 24-25 cm, and 40-43 cm, and in Samples 198-1210B-24H-1, 5-6 cm, and 24H-1, 48-49 cm. Sample 198-1210B-24H-1, 5-6 cm, yields abundant woodringinids, which outnumber the low trochospiral group, including P. eugubina. The underlying sample (198-1210B-24H-1, 36-37 cm) is characterized by common larger P. eugubina with subordinate biserial heterohelicids. Few biserial woodringinids and chiloguembelinids in association with common Guembelitria and rare P. eugubina are present in Sample 198-1210B-24H-1, 48-49 cm.
) is present in Samples 198-1210A-24H-4, 24-25 cm, and 40-43 cm, and in Samples 198-1210B-24H-1, 5-6 cm, and 24H-1, 48-49 cm. Sample 198-1210B-24H-1, 5-6 cm, yields abundant woodringinids, which outnumber the low trochospiral group, including P. eugubina. The underlying sample (198-1210B-24H-1, 36-37 cm) is characterized by common larger P. eugubina with subordinate biserial heterohelicids. Few biserial woodringinids and chiloguembelinids in association with common Guembelitria and rare P. eugubina are present in Sample 198-1210B-24H-1, 48-49 cm.
Cretaceous
The highest occurrence of Cretaceous planktonic foraminifers is in Samples 198-1210A-24H-4, 49-50 cm, and 198-1210B-24H-1, 57-58 cm. This assemblage is extremely dissolved with few whole specimens. Fragments of Cretaceous specimens contribute >50% of the fine fraction (38-150 µm), with the remaining part of the sample consisting of well-preserved tiny planktonic foraminiferal specimens that include Guembelitria and rare Hedbergella holmdelensis. Diverse assemblages of moderately to well-preserved planktonic foraminifers are present throughout the thick Campanian-Maastrichtian section cored at Site 1210. The Abathomphalus mayaroensis Zone (KS31) extends from the K/T boundary downward through Cores 198-1210A-27H and 198-1210B-25H, based on the presence of the nominal taxon. Additional taxa include A. intermedius, Contusotruncana contusa, Globotruncana linneiana, G. orientalis, G. rosetta, Globotruncanella havanensis, G. petaloidea, Globotruncanita stuarti, G. stuartiformis, Heterohelix globulosa, H. rajagopalani, Pseudoguembelina costulata, P. excolata, Pseudotextularia elegans, P. nuttalli, Racemiguembelina fructicosa, Rugoglobigerina hexacamerata, R. rugosa, and Planoglobulina multicamerata.
The FOs of R. fructicosa and C. contusa in Sample 198-1210B-27H-CC define a recognizable interval in the upper part of Zone KS30. The base of the Gansserina gansseri Zone (Zone KS30) is present in Sample 198-1210B-32H-CC. In addition to many of the taxa found in the overlying A. mayaroensis Zone, other characteristic species found throughout Zone KS30 include the nominal taxon, Contusotruncana fornicata, Globotruncana aegyptiaca, G. arca, G. bulloides, and G. falsostuarti. The base of the Globotruncana aegyptiaca Zone (Zone KS29) is found in Sample 198-1210B-33H-CC. The total range of Radotruncana calcarata defines the base of the Globotruncanella havanensis Zone (Zone KS28) in Sample 198-1210B-35H-CC and the base of the R. calcarata Zone (Zone KS27) in Sample 198-1210B-39H-CC. The Globotruncana ventricosa Zone (KS26) is present to the bottom of the cored section at Site 1210.
Benthic foraminifers were examined in selected core catcher samples from Holes 1210A and 1201B (Tables T5, T6). They are generally well preserved and rare to few in abundance in the Pleistocene through the Oligocene. The Eocene through Cretaceous section contains moderately preserved benthic foraminifers that are generally rare to few in abundance. The dissolution of planktonic foraminifers resulted in an increased relative abundance of benthic foraminifers in the upper to middle Miocene, in the Eocene/Oligocene boundary interval, and near the K/T boundary.
Neogene
In Samples 198-1210A-1H-CC through 10H-CC (Pleistocene-Pliocene), benthic assemblages are represented by abundant Oridorsalis tener, cibicidoidids (Cibicidoides wuellerstorfi and C. mundulus), uvigerinids (Uvigerina hispida, U. hispidocostata, U. senticosa, and U. peregina), Pyrgo murrhina, P. lucernula, and several agglutinated taxa (Eggerella bradyi, Karreriella bradyi, and Martinottiella sp.). In Samples 198-1210A-10H-CC to 12H-CC, stilostomellids (Stilostomella abyssorum, S. subspinosa, S. gracillima, and Stilostomella spp.) replace the uvigerinids. These Miocene benthic assemblages are also characterized by Oridorsalis tener and nodosariids (Dentalina spp. and Chrysalogonium longicostatum).
Paleogene
Beneath the Oligocene-Miocene condensed interval or unconformity, Sample 198-1210A-13H-CC contains abundant Bulimina jarvisi, Globocassidulina subglobosa, and Stilostomella subspinosa. Other components of the benthic assemblage include Oridorsalis umbonatus, buliminids (B. elongata and B. impendens), and Vulvulina spinosa. Upper Eocene Sample 198-1210A-14H-CC contains well-preserved and abundant benthic foraminifers. This assemblage is dominated by very large specimens of Plectofrondicularia paucicostata. Nuttallides truempyi, O. umbonatus, some cibicidoidids (Cibicidoides eocaenus and C. havanensis), Dentalina spp., Bulimina jarvisi, and V. spinosa are also present in this sample. In Sample 198-1210A-16H-CC, species such as Alabamina dissonata, N. truempyi, Dentalina spp., buliminids (B. impendens, B. jarvisi, and B. semicostata), and stilostomellids (Stilostomella spp. and S. gracillima) characterize the benthic assemblages.
The preservation and size of benthic foraminiferal tests decrease in lower Eocene (Samples 198-1210A-17H-CC through 19H-CC). Few benthic foraminifers are present in the >250-µm range. Small trochospiral forms (Nuttallides truempyi, Oridorsalis umbonatus, and Clinapertina complanata), Dentalina spp., abundant Bulimina jarvisi, and B. semicostata characterize benthic assemblages in this interval, although buliminids are absent in Sample 198-1210A-19H-CC.
The Paleocene benthic assemblages of Samples 198-1210A-20H-CC to 23H-CC and 198-1210B- 23H-CC contain various trochospiral forms (Conorbinoides hillebrandti, Gyroidinoides globosus, Nuttallides truempyi, Oridorsalis umbonatus, and Osangularia plummerae), Aragonia ouezzanensis, A. velascoensis, Lenticulina spp., and Buliminella grata. Although sporadic in their distribution, agglutinated taxa (Spiroplectammina jarvisi and Tritaxia spp.) are also present in this interval.
Cretaceous
Benthic assemblages in Samples 198-1210A-24H-CC to 26H-CC and 198-1210B-24H-CC to 32H-CC are represented by trochospiral species (Conorbinoides hillebrandti, Nuttallides truempyi, Oridorsalis umbonatus, and Sliterella lobulata), Dentalina spp., Lenticulina spp., and agglutinated taxa (Gaudryina pyramidata and Marssonella trochoides). Although they are sporadically present, Aragonia velascoensis and Pyramidina szajnochae are also characteristic taxa in this section.
In Samples 198-1210A-34H-CC through 42H-CC, Aragonia velascoensis, A. ouezzanensis, and the agglutinated species Gaudryina pyramidata and Marssonella trochoides are characteristic benthic foraminifers. Other important components of the benthic assemblages are similar to those observed in the upper part of the section.
Paleodepth
Paleodepth estimates are based on the work of Pflum et al. (1976), Tjalsma and Lohmann (1983), Woodruff (1985), and van Morkhoven et al. (1986) for the Neogene and Paleogene section. For the Cretaceous section, estimates are mainly based on the study of Nyong and Olsson (1984) and backtracked paleodepth-curve based on DSDP and ODP data from Kaiho (1999).
From the Neogene through Cretaceous, the observed benthic assemblages at Site 1210 are generally similar to those from Sites 1207-1209. Neogene paleowater depth is estimated to be upper abyssal (2000-3000 m) based on the presence of the characteristic forms as uvigerinids (Uvigerina hispida, U. hispidocostata, and U. senticosa), Pyrgo murrhina, P. lucernula, Eggerella bradyi, Karreriella bradyi, and Martinottiella sp. (Pflum et al., 1976; van Morkhoven et al., 1986).
Nuttallides truempyi, Oridorsalis umbonatus, Aragonia ouezzanensis, A. velascoensis, buliminids, and stilostomellids characterize Paleogene benthic assemblages. These also indicate water depths similar to those of the Neogene section (Tjalsma and Lohmann, 1983), but slightly shallower depths in the early Paleocene are suggested by the presence of such forms as Gyroidinoides globosus, Lenticulina spp., and buliminids (Tjalsma and Lohmann, 1983).
Trochospiral calcareous benthic foraminifers (Conorbinoides hillebrandti, Gyroidinoides globosus, Nuttallides truempyi, Oridorsalis umbonatus, Osangularia plummerae, and Sliterella lobulata) are characteristic taxa of Cretaceous benthic assemblages, and they indicate a wide bathymetric range (1500-4000 m) (Kaiho, 1998, 1999). Pyramidina szajnochae is found in Samples 198-1210B-28H-CC and 30H-CC, is one of the characteristic taxa of the Late Cretaceous, and is considered to indicate uppermost abyssal depth (2000-2500 m) (Nyong and Olsson, 1984). Furthermore, another dominant component, Aragonia velascoensis, which is indicative of lower bathyal depths (1500-2000 m) (Nyong and Olsson, 1984), becomes more dominant during the Campanian. In summary, this site shows a deepening upward trend from lower bathyal depths during the Campanian to upper abyssal depths during the Cenozoic.
