BIOSTRATIGRAPHY
Sediments of Site 1212 range in age from late Pleistocene to late Albian. Calcareous nannofossils are generally abundant and moderately to well preserved throughout the sediments of Holes 1212A and 1212B. Planktonic foraminifers are more variable in abundance and preservation, with the poorest preservation (high levels of test fragmentation) in the upper Miocene-Pleistocene. Sedimentation was not continuous and four unconformities/condensed intervals have been recognized separating upper Miocene/middle Miocene, middle Miocene/middle Eocene, mid-Campanian/lower Santonian-Coniacian, and Santonian-Coniacian/basal Cenomanian-uppermost Albian (see "Sedimentation and Accumulation Rates"). All core catcher samples were examined, and supplementary samples were used to refine datum positions and the biostratigraphy in and around critical intervals. Several critical intervals were identified from Holes 1212A and 1212B and include the PETM, the K/T boundary, and the MME.
Benthic foraminifers were examined in selected core catcher samples in Holes 1212A and 1212B. They are generally well preserved and rare to few in abundance in the Neogene through Cretaceous sections except for the upper Paleocene and middle Eocene sections, where relative benthic abundance is high because of planktonic foraminiferal dissolution. Paleodepth estimates are based on the work of Pflum et al. (1976), Tjalsma and Lohmann (1983), Woodruff (1985), and van Morkhoven et al. (1986) for the Neogene and Paleogene section. For the Cretaceous section, estimates are mainly based on the study of Nyong and Olsson (1984) and the backtracked paleodepth curve from the DSDP and Ocean Drilling Program (ODP) data of Kaiho (1999).
Neogene
The Neogene section ranges from the upper Pleistocene (Zone CN15) to middle Miocene (Zones CN3 and CN4). The Pleistocene to upper Miocene interval appears to be relatively complete and exhibits a high sedimentation rate that is typical of other Neogene sections recovered during Leg 198 (Table T2). In mid- to lower Pliocene samples from Cores 198-1212A-4H through 7H (e.g., Samples 198-1212A-4H-CC and 5H-1, 45 cm) numerous reworked taxa of earliest Pliocene and late Miocene age, including Reticulofenestra pseudoumbilicus, Amaurolithus spp., and Discoaster quinqueramus, have been recognized. Care must be taken with the LOs of these species within this interval.
An unconformity that includes most of the upper and middle Miocene is constrained by the FO of Discoaster berggrenii and the LO of Sphenolithus heteromorphus (Samples 198-1212A-8H-3, 5 cm, to 110 cm, and 198-1212B-7H-6, 5 cm, to 120 cm). Another significant unconformity separating lower middle Miocene and middle Eocene sediments lies just below this interval and is constrained by the occurrence of Sphenolithus heteromorphus (Zones CN3 and CN4) above and the occurrence of Discoaster sublodoensis (Zones CP12-lower CP13) below.
Paleogene
Paleogene sediments range from middle Eocene (Zone CP12) to earliest Paleocene (Subzone CP1a) in age, based on the FO of Discoaster sublodoensis and the FO of Biantholithus sparsus, respectively. Sedimentation rates are lower than in the overlying Neogene (see "Sedimentation and Accumulation Rates").
The P/E boundary and PETM interval appear to be relatively complete based on the levels of several datums. These datums include the LO of Fasciculithus (Samples 198-1212A-10H-1, 1 cm, and 198-1212B-9H-5, 55 cm), which lies just above the clay horizon that marks the PETM in the upper part of Zone CP8, the FO of Discoaster diastypus (Sample 198-1212A-9H-7, 30 cm), and the FO of Tribrachiatus orthostylus at the base of Subzone CP9a (Sample 198-1212A-9H-1, 10 cm). Not all datums were determined in both cores, due to sampling of inadequate resolution.
The K/T boundary is present in Cores 198-1212A-12H and 198-1212B-11H and appears to be complete. This interval follows the typical succession observed at other Leg 198 sites and includes the LO of Micula prinsii at the top of Zone CC26 and the presence of bladelike calcite below the boundary and the presence of common calcispheres above the boundary (Samples 198-1212A-12H-7, 73 cm, and 198-1212B-11H-7, 20 cm). These sediments are immediately overlain by an interval devoid of nannofossils, except for a few "survivor" species, such as Markalius inversus and Goniolithus fluckigeri, and reworked Cretaceous taxa (Samples 198-1212A-12H-7, 60 cm, and 198-1212B-11H-7, 3 cm). This interval is capped by sediments of Subzone CP1a, including taxa such as Biantholithus sparsus and Neobiscutum spp. (Samples 198-1212A-12H-6, 80 cm, 198-1212B-11H-4, 70 cm, 198-1212A-12H-7, 1 cm, and 198-1212B-11H-6, 70 cm, respectively).
Cretaceous
Sediments of late Maastrichtian to late Albian age were recovered from Holes 1212A and 1212B. High sedimentation rates characterize the Maastrichtian and upper Campanian interval. The nannofossil assemblages from these sediments are typically well preserved and diverse. The Maastrichtian appears to be relatively complete. Sediments assigned to Zone CC22 (Campanian) overlie a short section of Coniacian-aged chalk in Section 198-1212B-24H-6, based on the presence of Lithastrinus septenarius and the absence of Micula staurophora (Sample 198-1212B-24H-6, 30 cm). This, in turn, overlies sediment assigned to lower Subzone NC10a (basal Cenomanian-upper Albian) based on the presence of Eiffellithus turriseiffelii and Broinsonia stenostaurion (interval 198-1212B-24H-6, 40-51 cm).
Neogene
The upper 60 m of section at Site 1212 records continuous deposition of uppermost Miocene-Pleistocene sediment based on the recognition of planktonic foraminiferal Zones N18-N22 (Table T3). Preservation of planktonic foraminifers varies from poor to moderate, with selective dissolution and fragmentation leading to the sporadic presence of some biostratigraphically useful taxa through this interval. Most of the core catcher samples examined from Holes 1212A and 1212B are dominated by dissolution-resistant taxa including Globorotalia inflata (upper Pliocene-Pleistocene), Truncorotalia crassaformis, Sphaeroidinella dehiscens, Sphaeroidinellopsis seminulina (uppermost Miocene-lower Pliocene), and Sphaeroidinellopsis subdehiscens, with persistent but lower numbers of Globigerina bulloides, Neogloboquadrina pachyderma (dextral), Neogloboquadrina dutertrei, Globorotalia tumida, Globoconella conomiozea (lower Pliocene), Globigerinoides ruber, Globigerinoides sacculifer, G. obliquus, and Orbulina universa. Occasional downhole contamination in the core catcher samples is suspected based on the presence of very rare Truncorotalia truncatulinoides below its predicted range mixed with specimens indicative of Zone N19 in Sample 198-1212A-4H-CC. In addition, reworking of older sediments or downhole mixing of assemblages is responsible for the co-occurrence of Truncorotalia tosaensis (upper Pliocene-Pleistocene) and Globorotalia margaritae (uppermost Miocene-lower Pliocene) in Sections 198-1212A-4H-CC and 5H-CC. Section 198-1212A-7H-CC contains early forms of G. tumida with G. margaritae, Globorotalia plesiotumida, and Globigerinoides conglobatus, without Sphaersoidinella dehiscens, suggesting a Zone N18 assignment.
An unconformity or condensed section in Sections 198-1212A-8H-3 and 198-1212B-7H-6 separates the uppermost Miocene (Zone N18 or N17) and lower middle Miocene (Zone N9). Sample 198-1212B-7H-6, 118-120 cm, was collected near the base of this dark-colored interval and contains Orbulina universa, Praeorbulina glomerosa, Paragloborotalia mayeri, Globigerinoides mitra, Sphaeroidinellopsis seminulina, Sphaeroidinellopsis disjuncta, and Fohsella peripheroronda.
Paleogene
The lower middle Miocene rests unconformably on lower middle Eocene sediments assigned to Zone P11 based on the presence of Morozovella aragonensis, Igorina broedermanni, Acarinina pentacamerata, Truncorotaloides libyaensis, and Globigerinatheka senni in Sample 198-1212B-7H-6, 135-137 cm. A relatively thin Paleocene-lower middle Eocene section was cored at Site 1212, and many of the planktonic foraminiferal zones were not observed during the course of core catcher examination because of the low sedimentation rates (see "Sedimentation and Accumulation Rates"). Planktonic foraminiferal preservation varies from moderate to good.
The LO of Morozovella velascoensis, which lies a short distance above the onset of the PETM, lies between Samples 198-1212B-9H-4, 114 cm, and 9H-5, 42 cm. The base of the PETM event, however, appears to correspond to an unconformity in Hole 1212B. Foraminifers in Sample 198-1212B-9H-5, 77-78 cm, include Globanomalina pseudomenardii, the LO of which defines the top of Zone P4 at 55.9 Ma, some 0.4 m.y. before the onset of the event.
A paleontologically complete K/T boundary section was recovered at Site 1212. The boundary is bioturbated, and burrows of the lowermost Danian sediment extend 10 cm into the uppermost Maastrichtian. These lowermost Danian sediments yield a tiny Guembelitria assemblage reminiscent of the P0 zonal fauna in the deepest burrows, followed above by the assemblage of Zone P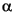 , defined by the first occurrence of Parvularugoglobigerina eugubina, in Samples 198-1212A-12H-7, 23-24 cm, 12H-7, 72 cm, and 198-1212B-11H-7, 23-24 cm. The former sample contains rare ~100-µm-sized, amber to olive-green spherules, probably altered tektites. The spherules are concentrated in the first 2-3 cm above the K/T boundary within Zone P. At Site 1212, in the lowest 10-20 cm of the basal Paleocene, planktonic foraminifers are extremely abundant representing up to 34% of smear slides.
, defined by the first occurrence of Parvularugoglobigerina eugubina, in Samples 198-1212A-12H-7, 23-24 cm, 12H-7, 72 cm, and 198-1212B-11H-7, 23-24 cm. The former sample contains rare ~100-µm-sized, amber to olive-green spherules, probably altered tektites. The spherules are concentrated in the first 2-3 cm above the K/T boundary within Zone P. At Site 1212, in the lowest 10-20 cm of the basal Paleocene, planktonic foraminifers are extremely abundant representing up to 34% of smear slides.
Cretaceous
A relatively thick and continuous section of mid-Campanian-Maastrichtian nannofossil ooze was recovered in Hole 1212B based on the complete succession of planktonic foraminiferal zones from the Radotruncana calcarata (Zone KS27) through the Abathomphalus mayaroensis (Zone KS31) Zones in intervals 198-1212A-12H-CC through 13H-CC and 198-1212B-11H-CC through 23H-CC (Table T3). The ooze immediately underlying the K/T boundary correlates to the uppermost Maastrichtian Abathomphalus mayaroensis Zone (Zone KS31) based on the presence of the nominal taxon, but the planktonic foraminiferal assemblages are affected significantly by dissolution. Below this level, however, assemblages are diverse and generally well preserved. Section 198-1212B-24H-6 contains a 20- to 25-cm-thick section of intact chalk containing planktonic foraminifers of Coniacian-early Santonian age, thereby suggesting an unconformity in the highly disturbed sediments of Core 198-1212B-24H.
Another stratigraphic break separating the Coniacian-lower Santonian and basal Cenomanian is present between Cores 198-1212B-24H and 25H. Sample 198-1212B-25H-1, 42-43 cm, contains planktonic foraminifers indicative of the lower Cenomanian-uppermost Albian Rotalipora globotruncanoides Zone (Zone KS17) based on the presence of the nominal taxon without Planomalina buxtorfi. Samples 198-1212B-25H-CC and 27H-CC are assigned to the upper Albian Rotalipora appenninica Zone (Zone KS16) based on the presence of Hedbergella libyca in the former sample and the co-occurrence of rare R. appenninica and P. buxtorfi in the latter sample.
Neogene
Samples 198-1212A-1H-CC through 5H-CC (Pleistocene-lower Pliocene), benthic assemblages are characterized by Cibicidoides wuellerstorfi, Oridorsalis tener, Pullenia bulloides, Pyrgo murrhina, and uvigerinids (Uvigerina hispida and Uvigerina hispidocostata). Samples 198-1212A-6H-CC and 7H-CC (lower Pliocene-upper Miocene) are dominated by stilostomellids (Stilostomella abyssorum, Stilostomella gracillima, and Stilostomella subspinosa). Additional taxa include O. tener, P. bulloides, P. murrhina, and agglutinated taxa (Karreriella bradyi and Martinottiella sp.) (Table T4).
These characteristic benthic assemblages indicate upper abyssal depths (2000-3000 m), even though taxa such as Pullenia bulloides show a wide bathymetric range from lower neritic-abyssal depth (Pflum et al., 1976).
Paleogene
In Samples 198-1212A-8H-CC (middle Eocene) and 9H-CC (uppermost Paleocene), benthic foraminifers are very rare in abundance and moderately preserved. Their test sizes are small, and most of the specimens are found in the 125- to 250-µm size fraction, except for nodosariids (Dentalina spp. and Nodosaria spp.). Benthic assemblages in Sample 198-1212A-8H-CC are characterized by small-sized buliminids (Buliminella grata, Bulimina impendens, and Bulimina jarvisi), Clinapertina complanata, Dentalina spp., and Nuttallides truempyi. In Sample 198-1212A-9H-CC, N. truempyi, Oridorsalis umbonatus, and Quadrimorphina profunda characterize the assemblage. No agglutinated forms are present in these two samples.
In Samples 198-1212A-10H-CC through 12H-CC and 198-1212B-10H-CC, benthic assemblages are generally represented by abundant Oridorsalis umbonatus, Gyroidinoides globosus, Nuttallides truempyi, Dentalina spp., Lenticulina spp., abundant Tritaxia (Tritaxia globulifera, Tritaxia spp., and Tritaxia rugolosa), and Spiroplectammina jarvisi. Though rare in abundance, Aragonia ouezzanensis, Aragonia velascoensis, and buliminids (Bulimina trinitatensis, Bulimina velascoensis, Buliminella grata, and Quadratobuliminella pyramidalis) are also characteristic forms in these sections (Table T4). These Paleogene benthic assemblages are indicative of upper abyssal depths (2000-3000 m), according to Tjalsma and Lohmann (1983) and van Morkhoven et al. (1986).
Cretaceous
The Upper Cretaceous Samples 198-1212A-13H-CC and 198-1212B-12H-CC through 23H-CC contain abundant O. umbonatus, G. globosus, N. truempyi, Nuttallinella florealis, Aragonia ouezzanensis, A. velascoensis, Dentalina spp., Lenticulina spp., and abundant agglutinated taxa (Marssonella trochoides, Gaudryina pyramidata, Spiroplectammina sp., S. jarvisi, Tritaxia spp., T. globulifera, and T. rugolosa). Section 198-1212B-24H-CC is represented by highly disturbed sediment that contains an assemblage with Albian-Cenomanian benthic foraminifers, including Conorotalites aptiensis and Hanzawaia compressa, mixed with younger Cretaceous taxa.
Samples 198-1212B-25H-CC and 27H-CC are characterized by Lower Cretaceous benthic foraminifers including abundant trochospiral calcareous species (Conorotalites aptiensis, Gyroidinoides globosus, Gyroidinoides infracretaceus, Hanzawaia compressa, Protosangularia albiana, and Protosangularia cenomaniana), diverse nodosariids (Astacolus spp., Dentalina spp., Lenticulina spp., Marginulina inaequalis, Nodosaria spp., and Saracenaria triangularis), and abundant agglutinated taxa (Dorothia gradata, Marssonella oxycona, Pseudoclavulina sp., Rhizammina spp., Spiroplectammina sp., Spiroplectinella excolata, Tritaxia spp., and T. globulifera). Pleurostomella spp. is also abundant in these samples (Table T5).
