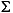 H2S = H2S + HS-) is below the detection limit (0.0002 mM) throughout the entire sediment column.
H2S = H2S + HS-) is below the detection limit (0.0002 mM) throughout the entire sediment column.
Site 1231 was selected for drilling in order to study the microbial activities and communities of the organic-poor sediments that characterize much of the world's open ocean. Before drilling at Site 1231, the nature of subseafloor microbial communities in open-ocean clays had never been assessed.
The principal objectives at this site were
Site 1231 is in the Peru Basin at 4827 m water depth. The lithologies, age, and many geochemical characteristics of the targeted sediments were characterized by Leg 34 studies at nearby Site 321 (Shipboard Scientific Party, 1976). The total sediment thickness at Site 321 is 115 m. The sediment is composed of 58 m of late Oligocene to Holocene clay and 57 m of iron-rich late Eocene to early Oligocene nannofossil ooze (Shipboard Scientific Party, 1976). The lower 50 m of sediment at Site 321 is rich in iron and manganese (Dymond et al., 1976). Iron and manganese accumulation rates estimated for the sediments present below 49 mbsf are about an order of magnitude higher than those estimated for sediments above 49 mbsf (Boström et al., 1976). In analyses of six interstitial water samples, dissolved manganese was present at relatively higher concentrations in the upper 45 m of the sediment column (3.5-7.4 ppm) than in the lower 50 m (0-1 ppm) (Brady and Gieskes, 1976). Dissolved sulfate concentrations also appeared to be slightly higher in the upper 45 m (>28 mM) than in the lower 50 m (27 mM) (Brady and Gieskes, 1976). Little or no evidence for other postdepositional reactions was seen among major dissolved ions at Site 321. This finding led Brady and Gieskes (1976) to conclude that any reactions in these sediments occur at such slow rates that their chemical signature is annihilated by diffusional exchange with the top and bottom of the sediment column. Consequently, Site 1231 provided a challenging opportunity for assessing the microbial activities and communities of low-activity sediments typical of much of the open ocean.
The subsurface extent of key electron donors (hydrogen, acetate, and formate), electron acceptors with standard free-energy yields greater than that of sulfate (oxygen and nitrate), products of key metabolic reactions (dissolved iron), and other biologically important chemicals was not determined for Site 321.
At Site 1231, the DIC profile hovers slightly at or below 3.3 mM for most of the sediment column. It exhibits three slight exceptions to this relative constancy: it slightly increases from 2.8 mM near the sediment/water interface, it exhibits a small peak of ~3.7 mM centered at 55 mbsf, and it declines slightly to ~3.0 mM at the sediment/basement interface. These DIC concentrations are even lower than those at Site 1225 (3.0-4.0 mM). They are much lower than the DIC concentrations observed at the other Leg 201 sites. Dissolved ammonium concentrations are also generally lower at Site 1231 than at the other Leg 201 sites. As at Site 1225, concentrations of DIC, ammonium, and alkalinity peak in the middle of the sediment column and decline toward both the sediment/ocean interface and the sediment/basement interface. The relatively low variability in the concentration profiles of these chemical species suggests that net microbial activity is lower at Site 1231 than at any other Leg 201 site. The midcolumn peaks in these profiles and their relatively low values near both the sediment/water and sediment/basement interfaces indicate chemical exchange from the sediment to the ocean and from the sediment to the basement.
The dissolved sulfate concentrations are >28 mM at the sediment surface and decrease linearly to 27 mM near the basement. The slight total downhole decrease in sulfate concentrations suggests that Site 1231 is characterized by lower sulfate-reducing activity than all of the other Leg 201 sites. Dissolved sulfide (H2S = H2S + HS-) is below the detection limit (0.0002 mM) throughout the entire sediment column.
Electron acceptors with higher standard free-energy yields than sulfate are present throughout most of the sediment column at Site 1231. Dissolved nitrate appears to be present in the uppermost meter and the lowermost 60 m of the sediment column (where it ranges from 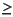 15 µM at 114 mbsf to 2 µM at 77 mbsf). Dissolved oxygen similarly appears to be present in the top 0.6 m below the seafloor as well as the last 3.8 m of sediment above basaltic basement. The diffusion of oxygen and nitrate from the overlying ocean down into the sediment is readily predictable from deep-ocean chemistry. However, the first Leg 201 locations, Sites 1225 and 1226, provided the only previous precedent for upward transport of nitrate (and, at Site 1225, oxygen) into deeply buried sediment from the underlying basaltic crust. As at Site 1225, the introduction of dissolved nitrate high into the sediment column at Site 1231 indicates that nitrate-utilizing microbial activity is present but may proceed at a very low rate in the site's lowermost sediments. Also as at Site 1225, the presence of dissolved oxygen and nitrate in these deepest sediments suggests that microbial activity in the underlying basalt is insufficient to strip even the scarcest preferentially utilized electron acceptors from the water that flows through the basalt.
15 µM at 114 mbsf to 2 µM at 77 mbsf). Dissolved oxygen similarly appears to be present in the top 0.6 m below the seafloor as well as the last 3.8 m of sediment above basaltic basement. The diffusion of oxygen and nitrate from the overlying ocean down into the sediment is readily predictable from deep-ocean chemistry. However, the first Leg 201 locations, Sites 1225 and 1226, provided the only previous precedent for upward transport of nitrate (and, at Site 1225, oxygen) into deeply buried sediment from the underlying basaltic crust. As at Site 1225, the introduction of dissolved nitrate high into the sediment column at Site 1231 indicates that nitrate-utilizing microbial activity is present but may proceed at a very low rate in the site's lowermost sediments. Also as at Site 1225, the presence of dissolved oxygen and nitrate in these deepest sediments suggests that microbial activity in the underlying basalt is insufficient to strip even the scarcest preferentially utilized electron acceptors from the water that flows through the basalt.
Dissolved manganese is present from 1 to 65 mbsf at Site 1231. Concentrations steadily rise from ~17 µM at 1.4 mbsf to a local peak of 78 µM at ~17 mbsf, decline briefly by a few micromolar, and then rise to sustain the highest concentration of 120 µM from 36 to 46 mbsf. Manganese concentrations below this peak steadily decline to essentially 0 µM at 68 mbsf. A relatively broad zone of generally high but variable dissolved iron concentrations (7-36 µM) spans the interval from 1 to 30 mbsf. A very small secondary peak in dissolved iron (5 µM) is centered near 74 mbsf. Two aspects of these broad patterns run counter to the general expectation that manganese reduction occurs at shallower depths than iron reduction in marine sediments. The first aspect is the broad co-occurrence of dissolved iron and manganese from 1 to 30 mbsf. The second aspect is the presence of the maximum dissolved iron concentration much closer than the maximum dissolved manganese concentration to the sediment/water interface. It appears likely that rates of manganese reduction in these sediments are limited by the availability of manganese oxides that supply dissolved manganese. Rates of iron reduction may be similarly limited by the presence and solubility of the minerals that supply dissolved iron.
As at other Leg 201 sites, the downhole distribution of microbial manganese and iron reduction at Site 1231 appears to be ultimately determined by lithology and depositional history. The peak intervals of dissolved manganese production are limited to the clays that lie between 11 and 55 mbsf. The maximum manganese concentration (120 µM) is present in the yellow volcanic-rich clay of Subunit IIA (31-44 mbsf). The secondary peak (78 µM) is centered in the green diatom-rich clay of Unit I (11-30 mbsf). Dissolved iron is similarly limited to the clay-rich portions of the upper sediment column. It exhibits a sharp maximum concentration (36 µM) a few meters below the seafloor in the radiolarian- and clay-rich diatom ooze of upper Unit I (0-11 mbsf). Most of the dissolved iron at Site 1231 is present in a broad maximum of 26 µM in the green clay of Unit I. Dissolved iron concentrations are consistently 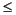 5 µM in the nannofossil oozes that lie between 55 mbsf and the basaltic crust (114 mbsf). Dissolved manganese is consistently <1 µM over the same interval.
5 µM in the nannofossil oozes that lie between 55 mbsf and the basaltic crust (114 mbsf). Dissolved manganese is consistently <1 µM over the same interval.
Although Site 1231 may be the microbially least active of the Leg 201 sites, its sediments still contain methane at concentrations of up to 15 µM. At this site, methane is limited to the upper clay-rich portion of the sediment column between 0 and 42 mbsf. This methane-bearing interval is completely within the interval of high dissolved manganese concentrations. Interestingly, this methane was only detected after prolonged incubation of headspace samples over a couple of days, whereas short 20-min incubation according to the standard ODP safety protocol showed only trace methane concentrations throughout the sediment column. The appearance of methane over time is currently interpreted as a release of sorbed methane. From sediments below 42 mbsf, no release of sorbed methane was observed and the concentrations remained at trace levels of <0.2 µM. The relationship of this sorbed methane to current microbial activity remains unknown.
Acetate concentrations range between 1 and 14 µM at Site 1231. Formate varies between 1 and 19 µM. Concentrations of both fatty acids are lowest in the top 3 m below seafloor (1-2 µM). They are slightly higher (3-6 µM) in the nitrate-reducing zone that spans the last 50 m above basement. Acetate and formate exhibit their highest concentrations (4-14 µM and 9-19 µM, respectively) at intermediate sediment depths (25-75 mbsf and 25-80 mbsf, respectively). These broad patterns suggest that at Site 1231 fatty acid concentrations may be lower in the sedimentary intervals that include electron acceptors with the highest energy yields. Curiously, the acetate and formate concentrations at this site are generally an order of magnitude higher than concentrations in sediments of the equatorial Pacific sites but are similar to those found at the Peru margin sites and in other very active coastal marine sediments. As noted in the site chapters, accurate understanding of the fatty acid distribution and its microbial relevance will require thorough postcruise analyses of microbial energetics in subseafloor environments.
Hydrogen concentrations are extremely high in the uppermost 35 m of the sediment column, with a peak value of 102 nM at 15 mbsf. This is the highest hydrogen concentration measured at any Leg 201 site. It exceeds the hydrogen concentration of near-surface iron-reducing sedimentary environments by >100-fold. The zone of high hydrogen coincides with the zone of iron reduction but does not show any direct correlation with distributions of fatty acids or methane. The presence of extremely high hydrogen concentrations at the site with the lowest organic carbon mineralization rates remains unexplained at this point. From 44 mbsf down to the basaltic basement, hydrogen concentrations are, in contrast, very low (0.05-0.22 nM).
Prokaryotic cell counts were conducted on samples from throughout the sediment column at Site 1231. These data show that mean cell concentrations are generally lower at this open-ocean site than at any previously enumerated ocean drilling site. Cell concentrations exhibit a distinct local concentration peak at 10-15 mbsf, the approximate depth of the Unit I zone of iron and manganese reduction.
Experiments on major microbial processes and on enumeration of viable prokaryotes were initiated at selected depths ranging from near the seafloor to the bottom of the drilled sediment column. The studied processes include methane and acetate formation and consumption, sulfate reduction, hydrogen oxidation, and rates of cell growth. The cultivation experiments include selective growth conditions for a wide range of autotrophic and heterotrophic prokaryotes ranging from psychrophilic to thermophilic. Cultivation experiments particularly focused on manganese- and iron-reducing bacteria throughout the column. Studies of sulfate-reducing bacteria in macrofaunal burrows were also initiated. Detailed microbiological sampling targeted sediment depths of particular biogeochemical interest, such as the midcolumn reduced manganese interface and the sediment/basalt interface.
The results from six Adara tool deployments define a temperature profile composed of two distinct intervals: a linear gradient of 90°C/km from 0 to 55 mbsf and a linear gradient of 35°C/km from 55 to 115 mbsf. The sediment/water interface temperature measured by a mudline Adara tool deployment is 1.7°C. The estimated temperature at the base of the drilled sediment column (115 mbsf) is 8.6°C. Throughout the entire drilled interval (0-121 mbsf), temperatures are in the psychrophilic range.
Trials were undertaken of two experimental tools at this site: the Davis-Villinger Temperature-Pressure Probe (DVTP-P) and the catwalk infrared (IR) camera. The single DVTP-P deployment indicated minor overpressure at 108 mbsf.
Sites 1225 and 1226 were selected as drilling targets because their microbial activities and cell counts were expected to be far below those in ocean-margin settings but above those in the lowest-activity open-ocean environments.
The principal objectives at Site 1225 were
Site 1225 is located in the eastern equatorial Pacific near the present-day boundary between the South Equatorial Current and the North Equatorial Countercurrent at 3760 m water depth. It lies in the sedimentary bulge created by the rain of biogenic debris from the relatively high productivity equatorial ocean. Geochemical studies of DSDP and ODP sites throughout this region have shown that seawater flows through the basaltic basement that underlies the sediments throughout this region (Baker et al., 1991; Oyun et al., 1995). Anomalously low conductive heat flow occurs throughout the region (Von Herzen and Uyeda, 1963; Sclater et al., 1976), possibly because the large-scale flow of relatively cool seawater through the basalts depresses conductive heat flow (Oyun et al., 1995).
The lithologies, sediment age, and many geophysical characteristics of the target site were well characterized by earlier studies of nearby Site 851 (Mayer, Pisias, Janecek, et al., 1992; Pisias, Mayer, Janecek, Palmer-Julson, and van Andel, 1995). Those studies indicated that the site is representative of a large portion of the eastern equatorial Pacific region. The sediments of Site 851 have a continuous biostratigraphy with a minimal age of 11 Ma at the basaltic basement. The gross lithologic and physical properties of the carbonate and siliceous oozes and chalk at Site 851 are characteristic of sediments throughout the region (Mayer, Pisias, Janecek, et al., 1992). The interstitial water chemical profiles at Site 851 exhibit clear evidence of seawater flow through the underlying basalts (and perhaps the lower part of the sediment column) (Oyun et al., 1995; Spivack and You, 1997).
Cragg and Kemp (1995) documented the presence of prokaryotic cells and activity throughout the sediment column at Site 851. For the first few tens of meters below seafloor, counts of both total cells and dividing cells were low relative to counts from similar depths at sites from the Peru shelf and the Japan Sea (Cragg and Kemp, 1995). At greater depths, Site 851 cell counts approached the averaged values from all previously counted sites.
Leg 138 shipboard chemistry showed that concentrations of several dissolved chemical species (ammonium, strontium, and silica) and alkalinity peaked midway down the sediment column. In contrast, dissolved sulfate exhibited maximum values near the sediment/water interface and the basement/sediment interface (Mayer, Pisias, Janecek, et al., 1992). These patterns of sedimentary interstitial water concentration are inferred to result from low levels of biological activity throughout the sediment column, coupled with diffusive exchange with the overlying ocean and with water flowing through the underlying basalts (and perhaps the lower part of the sediment column) (Spivack and You, 1997). Geochemical modeling suggests that net microbial sulfate reduction in the upper half of the Site 851 sediment column corresponds to a respiration rate of 10-9 to 10-8 mol CO2/cm2/yr (D'Hondt et al., 2002). This rate of respiration is only the barest fraction of the rate of carbon dioxide reduction by photosynthesis in the overlying equatorial ocean (10-3 mol/cm2/yr) (D'Hondt et al., 2002). The subsurface distribution of electron acceptors with higher standard free-energy yields (oxygen, nitrate, manganese oxide, and iron oxides) in this region was not determined for Site 851.
At Site 1225, concentrations of methane, ammonium, DIC, and alkalinity peak in the middle of the sediment column and decline toward both the sediment/ocean interface and the sediment/basement interface. In contrast, sulfate concentrations are lowest in the middle and lower part of the sediment column and nitrate and dissolved oxygen are present only at the ocean and basement interfaces. These profiles result from the balance between net subsurface microbial activities and small net fluxes of biologically utilized chemicals across the ocean/sediment and sediment/basement interfaces.
Interstitial water data also document dissolved oxygen penetration into the top 2 m of the sediment column, an interval containing nitrate in the top 1.5 m of the sediment, a peak concentration of dissolved manganese at 3.6 mbsf, a broad zone of relatively high dissolved iron centered at ~25 mbsf, and sinks for reduced manganese and dissolved iron at 100 mbsf. Sulfate concentrations decrease downhole by only ~7% from local bottom-water values; most of this decrease occurs in the upper ~60 m. This vertically extended sequence of successive interstitial water chemical zones closely resembles the centimeter- to decimeter-scale sequence seen in nearshore sediments (with depth-dependent transitions from a zone of oxygen reduction to successive zones of nitrate, manganese oxide, iron oxide, and sulfate reduction). These data are consistent with the hypothesis that subseafloor microbial communities preferentially utilize the available electron acceptor that yields the highest free energy of reaction.
In the lower portion of the sediment column, this vertical sequence of successive reduction zones is reversed as a result of water flow through the underlying basaltic basement. Diffusion of solutes from this basement water to the overlying sediment delivers nitrate to the lowermost 20 m of the sediment column (300 mbsf to basement) and possibly also dissolved oxygen to the lowermost meter of the column (319.3 mbsf to basement). This short interval of dissolved oxygen and nitrate is overlain by a broad zone of dissolved manganese centered near 250 mbsf and a broad peak of dissolved iron centered at ~230 mbsf. These dissolved nitrate and oxygen profiles show that electron acceptors yielding high free energies of reaction are introduced to at least some portions of the deep subseafloor biosphere by hydrologic processes. They also indicate that microbial activity in the underlying basalt is insufficient to strip even the scarcest preferentially utilized electron acceptors from the seawater that flows through the basalt at this site.
Dissolved hydrogen concentrations in incubations of Site 1225 sediments are generally in the range of 1-2 nM. Lovley and Goodwin (1988) and Hoehler et al. (2001) observed similar concentrations in experiments with near-surface aquatic sediments where sulfate reduction is the primary electron-accepting reaction. On the basis of their observations, Lovley and Goodwin (1988) hypothesized that hydrogen concentrations in aquatic environments are controlled by competition between different metabolic pathways. According to this hypothesis, prokaryotes using electron acceptors that yield higher free energies of reaction are able to operate at lower electron donor concentrations and thereby out-compete prokaryotes limited to electron acceptors that yield lower free energies of reaction. Documentation of these concentrations at Site 1225 suggests that even in low-activity subseafloor sediments, hydrogen concentrations may be controlled by the same thermodynamic competition between electron-accepting pathways as in high-activity sediments and can be predicted from the dominant pathway.
Methane is present at trace concentrations of <0.25 µM throughout the sediment column. This finding demonstrates the presence of methane in subseafloor sediments with sulfate concentrations that are very close to seawater values. The generation of methane in these sediments challenges models of microbial competition that are based on standard free energies. There are a number of possible reasons for the occurrence of methanogenesis in sulfate-rich sediments. For example, the methanogens and sulfate reducers may rely on different electron donors (e.g., the methanogens may utilize methylated amines and the sulfate reducers may rely on hydrogen and/or acetate) (Oremland and Polcin, 1982; Oremland et al., 1982b; King, 1984).
The steady-state maintenance of methane in the subseafloor sediments of Site 1225 indicates that if anaerobic methanotrophy occurs here, it does not drive methane concentrations below a few hundredths to tenths of micromolar. Concentrations are lowest near the sediment/ocean and sediment/basement interfaces, where methane may be oxidized by prokaryotes using electron acceptors that yield relatively high energies of reaction (such as nitrate or dissolved oxygen). The highest methane concentrations are present in the middle of the sediment column, where sulfate appears to be the principal terminal electron acceptor available. We hypothesize that the peak methane concentrations are held at the observed level (~0.15-0.25 µM) because sulfate-reducing methanotrophs cannot oxidize methane at lower concentrations under in situ conditions.
Concentrations of acetate and formate were <1 and <0.5 µM, respectively, throughout the sediment column. These concentrations are an order of magnitude lower than those measured in continental shelf sediments (Sørensen et al., 1981; Wellsbury and Parkes, 1995) and are also lower than in other deep sediment sites (Wellsbury et al., in press). These very low concentrations may be regulated by limiting energy yields or limited by the kinetics of active uptake by the anaerobic respiring prokaryotes. Since these results are among the first to demonstrate very low concentrations of short-chain fatty acids in cold, low-activity subsurface sediments, there is no database for comparison.
Comparison of Site 1225 physical property, sedimentology, and chemical records suggests that broad-scale patterns of past oceanographic change exert strong influence on present subseafloor metabolic activity. Concentrations of dissolved iron closely follow downhole variation in magnetic susceptibility and split-core reflectance, with peak concentrations of dissolved iron and solid-phase iron compounds (inferred from magnetic susceptibility) in the intervals from ~0 to 70 mbsf and 200 to 270 mbsf. The intact magnetic reversal record suggests that the magnetic phases were created during or shortly after sediment deposition. These intervening sediments are low in dissolved iron, are low in magnetic susceptibility, are characterized by the most intensely bioturbated intervals, and were deposited during a late Miocene-early Pliocene biogenic bloom that occurred throughout much of the global ocean (van Andel et al., 1975; Farrell et al., 1995; Dickens and Owen, 1999).
Four Adara tool deployments plus two deployments of the Davis-Villinger Temperature Probe (DVTP) defined a sediment/water interface temperature of 1.4°C and an estimated sediment/basement interface temperature of 7.0°C. The downhole temperature gradient curved slightly downward. The slight curvature appears to be best explained by a geologically recent decrease in basement temperature, perhaps a result of an increased rate of seawater flow through the basement. Throughout the sediment column, in situ temperatures were well within the range inhabited by psychrophilic prokaryotes.
Experiments on major microbial processes and experiments for enumeration of viable prokaryotes were initiated at selected depths ranging from near the mudline to near the basement, where samples were obtained within centimeters of the basalt. Subsamples for postcruise biomolecular assays and microbiological experiments were routinely taken from all of the distinct geochemical zones and lithologic subunits. Total cell numbers were enumerated on board. These cell counts are very close to data obtained from nearby Site 851 and consequently demonstrate the high reproducibility of AODC tests in subseafloor microbial studies.
At this site, novel experiments with core temperatures and contamination tracers were undertaken to determine how handling of cores and samples for microbiological studies might be improved. Catwalk experiments with an IR camera were used to assess the effects of different core handling procedures on transient warming of the core and, consequently, on the survival of temperature-sensitive prokaryotes.
Site 1226 was selected as a drilling target because its microbial activity was expected to be intermediate between that in ocean-margin settings and that in the lowest-activity open-ocean environments.
The principal objectives at this site were
Site 1226 (3297 m water depth) is located in the eastern equatorial Pacific, 300 km south of the Galapagos Islands, near the present-day boundary between the South Equatorial Current and the Peru Current. Near the sea surface in this region, the advection of water from the Peru Current results in relatively high nutrient levels and biological productivity (Chavez and Barber, 1987). According to its calculated backtrack path, this site has drifted eastward but has remained near its present latitude for most of its history (Pisias et al., 1995; Farrell et al., 1995). Sediment thickness at Site 1226 is 420 m. The oldest sediments immediately overlie basaltic basement and have a biostratigraphic age of 16.5 Ma (Shipboard Scientific Party, 1992a). As described in "Principal Results" in "Site 1225" geochemical studies of DSDP and ODP sites throughout this region have shown that seawater flows through the underlying basaltic basement (Baker et al., 1991).
The lithology, sediment age, and many geochemical and geophysical characteristics of the target site were well characterized by earlier studies of Site 846. The gross lithologic and physical properties of the carbonate and siliceous oozes and chalk at Site 846 are characteristic of sediments throughout the region (Shipboard Scientific Party, 1992a; Pisias, Mayer, Janecek, Palmer-Julson, and van Andel, 1995). Leg 138 studies showed that the region has undergone large variations in sediment accumulation over the course of its history. Accumulation of calcium carbonate and opal was unusually low at Site 846 during the Miocene "carbonate crash" of 11-7.5 Ma and was unusually high during the widespread Indo-Pacific "biogenic bloom" that occurred from ~7 to 4.5 Ma (Farrell et al., 1995). The organic carbon accumulation rate is presently high and appears to have gradually increased throughout the Pleistocene (Shipboard Scientific Party, 1992a; Emeis et al., 1995).
Leg 138 shipboard chemical studies of Site 846 show that concentrations of several dissolved chemical species (methane, ammonium, strontium, and silica) and alkalinity peak part way down the sediment column. In contrast, dissolved sulfate, lithium, and calcium exhibit maximum values near the sediment/water interface and the basement/sediment interface (Shipboard Scientific Party, 1992a).
As at Sites 851 and 1225, these patterns of sedimentary interstitial water concentration are inferred to result from modest levels of biological activity throughout the sediment column, coupled with diffusive exchange with the overlying ocean and with seawater flowing through the underlying basaltic basement. The sediments of Site 846 have a higher organic carbon content than the sediments of Sites 851 and 1225. Organic carbon content at Site 846 ranges from 0.2% to 1.0% and is highest in the Pleistocene and upper Pliocene deposits. Accordingly, Site 846 exhibits steeper gradients than Sites 851 and 1225 in interstitial water chemical species that respond to prokaryotic mineralization processes, such as sulfate, ammonium, and methane. The distinctly higher concentrations of methane at Site 846 than at Site 851 are particularly intriguing because methanogenesis is generally understood to be suppressed by sulfate-reducing bacteria and methane may be oxidized in the presence of sulfate.
The subsurface distribution of key electron donors (hydrogen, acetate, and formate) and of electron acceptors with higher standard free-energy yields (oxygen, nitrate, manganese oxide, and iron oxides) was not determined for Site 846.
Site 1226 provides an excellent series of samples from the sediment/water interface down to basement, including good cores from the contact zone between sediment and basalt. The geochemical gradients that span the 420-m-thick sediment column are bounded at the sediment/seawater interface and the sediment/basement interface by comparable but opposite reduction-oxidation (redox) zonations. Sulfate reduction appears to be the predominant electron-accepting pathway at this site. A broad maximum of DIC and ammonium in the interstitial water demonstrates the mineralization of organic material throughout the sediment column at several-fold higher rates than at Site 1225. Concentrations drop steeply to near-seawater values at the sediment/water interface and less steeply toward seawater values at the contact with basement. Water flow through the basement thus provides an effective sink for sedimentary metabolic DIC.
As at Site 1225, the overall chemical zonations are consistent with thermodynamic control of electron acceptor use by subsurface prokaryotes. The data show that water flow through the underlying basaltic basement introduces electron acceptors with high free-energy yields to sediment hundreds of meters below the seafloor. Relative to Site 1225, however, these zonations are more compressed because of higher rates of prokaryotic activities. Oxygen was not detected at any sediment depth within the column at Site 1226, and any oxic surface layer of sediment must thus have been closer to the ocean interface than the depth of our first dissolved oxygen measurement in Hole 1226B, in Section 201-1226B-1H-1, 10 cm. Nitrate, however, was detected in core sections nearest to the sediment/water and sediment/basement interfaces. As at Site 1225, nitrate diffuses upward into the overlying sediment from water flowing through the basement. However, at Site 1226 the diffusing nitrate barely penetrates into the sediment column before being reduced.
As the next electron acceptor in the classical redox sequence, the interstitial water distribution of manganese shows a more complex pattern. Dissolved manganese peaks just at the sediment/water interface and again 9 m below, followed by a steep drop to a zone of near-zero concentrations between 100 and 250 mbsf. Yet another distinct peak in manganese is present at 300 mbsf. At the bottom of the sediment column, manganese peaks again between 400 mbsf and the sediment/basement interface. The near-basement peak and the 300-mbsf peak together define a broad 160-m interval of unusually high dissolved manganese concentrations.
Comparison with the Leg 138 Initial Reports data indicates that the most deeply buried interval of high dissolved manganese is composed of hydrothermally influenced sediments immediately above the basement. The 300-mbsf peak is present in sediments that were deposited at low rates during the Miocene "carbonate crash." In contrast, the sediments that define the overlying interval of near-zero dissolved manganese concentrations were deposited at high rates during the 7- to 4.5-Ma "biogenic bloom" that occurred throughout much of the world ocean (Farrell et al., 1995). These results suggest that the availability of electron-accepting pathways to current subseafloor activity directly depends on broad-scale patterns of past oceanographic change. More detailed interpretation of these multiple zones of apparent manganese reduction and oxidation must await further solid-phase and interstitial water chemical analyses.
The zone of dissolved sulfide extends from just 5 m below the sediment/water interface to a depth of 280 mbsf, and the broad peak reaches 0.7 mM at ~100 mbsf. Throughout this sulfidic sediment column, iron is <10 µM but displays narrow peaks of ~40 µM just above and below the sulfide zone. This pattern reflects both the low equilibrium concentration of ferrous iron in sulfidic interstitial water and the presence of reducible iron near the sediment surface and in the deep sediment column, including a third iron peak at 380 mbsf. The interstitial water data identify a sink for both sulfide and manganese within an interface at 250-280 mbsf, where manganese may precipitate with sulfide.
Methane exhibits a broad peak at 100-250 mbsf with concentrations of 2-3 µM. Although this is still a trace level of biogenic methane, it is more than tenfold higher than at Site 1225. Sulfate is present at >80% of seawater concentration throughout the sediment column and indicates active sulfate reduction over the entire methane peak. The coexistence of methane and sulfate at these levels demonstrates the ability of methanogens to maintain an active metabolism in a high-sulfate environment where competition for energy substrates must be strong and where the methanogens may be limited to noncompetitive substrates (Oremland and Polcin, 1982; Oremland et al., 1982b). The results also show that sulfate-reducing bacteria in this environment are apparently unable to exploit methane beyond the existing low concentration.
Acetate and formate concentrations are low in the upper 0-100 m of sediment (<0.5 µM). At greater depths, their concentrations increase to 1-3 µM. This shift in concentrations appears to result from regulation mechanisms that are not yet understood for any sedimentary environment. The volatile fatty acids (VFAs), acetate and formate, are known to be important substrates for most anaerobic respiring bacteria and for methanogens (Winfrey and Ward, 1983; Wellsbury and Parkes, 1995). The interstitial water concentrations of these intermediate fermentation products are regulated by a balance between production and consumption. Concentrations of both acetate and formate are usually found to be relatively higher in organic-rich marine sediments, where they appear to be a function of the rate of fatty acid production and of the energy-yielding metabolism of the consumers. For example, sulfate reducers are able to outcompete methanogens in their efficiency of substrate uptake and thereby drive acetate and formate concentrations to lower levels. However, control of VFA concentrations by such competition is difficult to reconcile with their increased concentrations in the deeply buried Site 1226 sediments that exhibit high dissolved manganese and iron concentrations.
Hydrogen was very low in incubated sediment samples from Site 1226, ranging from 0.1 to 0.8 nM. This is below the equilibrium concentration of a few nanomolar measured in the sulfate reduction zone of more active nearshore sediments (Hoehler et al., 1998) and is even below the concentrations measured at Site 1225, where prokaryotic activity is significantly lower than at Site 1226. According to theoretical calculations of the minimum energy yield required for prokaryotic respiration (Thauer et al., 1977; Schink, 1997) and also according to hydrogen data from a range of sedimentary environments, equilibrium concentration of hydrogen is maintained at the lowest limit that provides the lowest required energy yield of the hydrogen-metabolizing prokaryotes (Hoehler et al., 2001). Based on the dissolved sulfate, manganese, and iron data, sulfate reduction is the predominant respiration process throughout most of the sediment, with the other electron acceptors gaining relative significance near the top and bottom of the sediment column. However, the Site 1226 hydrogen incubation concentrations are lower than those in surface sediments, where sulfate reduction is the predominant process. This finding suggests that either the Site 1226 sulfate-reducing communities derive the canonical minimum energy yield at lower hydrogen concentrations than surface sulfate-reducing communities or they utilize hydrogen at energy yields below the previously accepted theoretical limit.
Experiments on samples from selected sediment depths were conducted on the major microbial processes, including methanogenesis, acetogenesis, sulfate reduction, hydrogen oxidation, and prokaryotic growth. Although most of these data will be available only postcruise, initial results show a time constant of hydrogen turnover on the order of a few days. Other substrates for microbial activities will have much longer turnover of months to years, and only the postcruise radiotracer results will demonstrate these rates. Total prokaryotic cell counts show 106-107 cells in the upper 100 m of the sediment column. This is an order of magnitude higher than at Site 1225, in accordance with the higher availability of organic material at Site 1226. Below 100 mbsf, the cell concentrations are rather similar at the two sites. A broad spectrum of prokaryotic MPN counts and enrichments was initiated at this site, ranging from heterotrophs to autotrophs and from psychrophiles to thermophiles. Samples were also taken for cultivation from pieces of basaltic rock recovered at the bottom of Hole 1226B. Because of the slow growth rate of the indigenous microorganisms, successful counts and cultures will expectedly require many months to years for growth and development.
Contamination tests are very important for all the microbiological work and were done continuously throughout drilling by injecting PFT into the drilling water. In all cores that were used for microbiological experiments, counts, or isolations, a contamination test was also conducted with prokaryote-sized fluorescent microbeads released within the core catcher upon impact with the sediment (five tests at Site 1226). The detection limit of the PFT method is 0.02 µL drilling fluid (seawater)/g sediment. The results show low to nondetectable contamination in most piston (APC) cores (<0.1 µL drilling fluid/g sediment) but significant potential contamination in XCB cores where the sediment was also visibly disturbed. Subsampling of XCB cores was done here with a reduced sampling program from intact biscuits of sediment. Slurry samples used for an extensive program of microbiology and process studies all (apart from one) have nondetectable contamination when using the PFT method and nondetectable or extremely low contamination using the bead method.
Eight Adara tool deployments and four deployments of the DVTP define a sediment/water interface temperature of 1.7°C and an estimated sediment/basement interface temperature of 24.4°C. An accurate linear temperature gradient of 54°C/km was determined through the 420-m-thick deposit. As the sediment depth increases, temperatures thus shift from the psychrophilic microorganism range to the mesophilic range. Deployment of the corresponding pressure tool (DVTP-P) showed ambient hydrostatic pressure.
As at Site 1225, most cores from the first deep hole (Hole 1226B) were logged on the catwalk with an IR camera for postcruise analysis of the IR logging utility. In order to continue building a temperature database suitable for assessing the microbiological effectiveness of catwalk core handling strategies and for determining microbial cultivation strategies, the IR camera was also used to immediately log temperature gradients across cut section ends.
Background and Objectives. Site 1227 was one of three Leg 201 sites selected for drilling on the continental shelf of Peru. These shelf sites were collectively selected to provide records of microbial activities, communities, and geochemical consequences in organic-rich ocean-margin sediments.
The principal objectives at this site were
Site 1227 (427 m water depth) is in the immediate vicinity of Leg 112 Site 684, in a small fault-bounded sediment pond in the Trujillo Basin on the Peru continental shelf. The Trujillo Basin lies within the Peru upwelling zone, and its sediments are correspondingly rich in organic carbon. The TOC content of Site 684 sediment samples ranges between 1.2% and 10.6%, (Shipboard Scientific Party, 1988c). The average TOC concentration of these samples is approximately an order of magnitude higher than the average concentration at open-ocean Site 846 (Leg 201 Site 1226) (Shipboard Scientific Party, 1988c, 1992a). It is about two orders of magnitude higher than the TOC content of open-ocean Site 851 (Leg 201 Site 1225) (Shipboard Scientific Party, 1988c, 1992b).
Geochemical studies of Leg 112 sites show that brine is present below the seafloor in the Trujillo and Salaverry Basins (Suess, von Huene, et al., 1988). The composition of the brine differs from site to site, perhaps because of differences in its degree of dilution and the nature of its interaction with the surrounding sediments (Suess, von Huene, et al., 1988). Detailed chemical analyses indicate that this brine is of marine origin and is early Miocene in age (Kastner et al., 1990). The Leg 112 Initial Reports volume suggested that it enters the younger sediment column by diffusion from interstitial brine in underlying Miocene sediments (Suess, von Huene, et al., 1988). Kastner and colleagues (1990) inferred that it is emplaced by stratigraphically bounded advection from north to south. The sulfate depletion of the brine at Site 1227 presumably results from bacterial sulfate reduction closer to the brine's source (e.g., deeper in the sediment column). Whatever the brine's mode of emplacement, Site 1227 provides an opportunity to study how the presence of sulfate-depleted brine affects subseafloor life in organic-rich sediments. Consequently, it provides an excellent standard of comparison for Sites 1228 and 1229, which are affected by the intrusion of sulfate-rich brine into, respectively, sulfate-rich and sulfate-depleted sediments.
Leg 112 shipboard chemistry suggests that the concentrations of methane at Site 684 increase by at least three orders of magnitude (from 102 to 105 µL/L) over the first 50 to 60 mbsf and remain between 104 and 105 µL/L to at least 100 mbsf. Ethane and butane concentrations also increase downhole to maximum concentrations at ~60 mbsf (Shipboard Scientific Party, 1988c). In contrast, the concentrations of dissolved sulfate decline from a near-seawater value to zero over the uppermost 30 or 40 mbsf (Shipboard Scientific Party, 1988c). These profiles of dissolved hydrocarbons and sulfate indicate that the hydrocarbons and the sulfate are simultaneously destroyed by sulfate-reducing prokaryotic communities at ~40 mbsf.
Concentrations of several dissolved chemical species increase steadily to the base of the hole (ammonium, chloride, calcium, and magnesium). The increases in dissolved chloride, calcium, and magnesium provide evidence of the brine diffusing upward into the sediment column. Alkalinity exhibits a maximum value at ~40 mbsf, where the rate of anaerobic oxidation of methane appears to be greatest. The magnesium/calcium ratio peaks at 12 mbsf and steadily declines to the base of the hole, presumably as a result of dolomitization throughout the methane-rich sedimentary interval (Shipboard Scientific Party, 1988c).
All of these patterns of sedimentary interstitial water concentrations are inferred to result from relatively high levels of biological activity throughout the sediment column, coupled with diffusive exchange with the overlying ocean and with the brine introduced at depth. The subsurface extent of key electron donors (hydrogen, acetate, and formate) and electron acceptors with standard free-energy yields greater than that of sulfate (oxygen, nitrate, manganese oxide, and iron oxides) was not determined for Site 684.
Principal Results. Interstitial water studies at Site 1227 define one of the most highly resolved chemical records in ODP history. An important objective with these profiles is to identify and quantify zones of prokaryotic activity based on reactive interstitial water species. A deep brine dominates the profiles of conservative seawater ions at this site, including chloride, which increases (with a linear gradient of 5 mM/m down to 70 mbsf and of 3 mM/m below that) to reach twice seawater chlorinity at 120 mbsf. Downhole depletion of sulfate at a relatively shallow depth, DIC concentrations as high as 25 mM, ammonium rising to 23,000 µM at 150 mbsf, and very high concentrations of methane all indicate that prokaryotic activity is much higher at this ocean-margin site than at open-ocean Sites 1225 and 1226. Dissolved sulfate concentrations rapidly decline in the upper 15 mbsf from a seawater value of 29 mM to 5 mM. Concentrations then decline more slowly to 0 mM at ~40 mbsf. Concentrations of dissolved hydrogen sulfide rise rapidly over the same 0- to 40-mbsf interval, from 0.04 mM at 0.24 mbsf to 9 mM at 39-40 mbsf. The convex-upward shape of both the sulfate and sulfide profiles from the sediment/water interface to ~40 mbsf indicates that bacterial sulfate reduction occurs throughout the interval. Sulfide concentrations steadily decline over the sulfate-poor remainder of the drilled section to <0.3 mM at 150 mbsf.
From 1 to 31 mbsf, dissolved barium concentrations rise slightly, from 0 to 1.9 µM. Over the next several meters, barium concentrations rise at an increasingly steep rate, climbing from 9 µM at 38 mbsf to 170 µM at 43 mbsf. Concentrations then rise steadily to 350 µM at ~150 mbsf. Dissolved sulfate and barium are both present throughout the entire interval of non-zero sulfate. Throughout this interval, the concentrations of dissolved barium and dissolved sulfate appear to be related by the solubility product of barite. Upward diffusion of barium from 43 to 38 mbsf appears to sustain modern barite formation in this Peruvian shelf sediment. The barite is visible as lighter bands in the sediment column and was confirmed by X-ray diffraction. At slightly greater depth (~42 mbsf), dissolved sulfate concentrations decline toward 0 mM, barite begins to dissolve, and dissolved barium concentrations rise. The narrow barium peak centered at 43 mbsf is inferred to mark the principal depth of current barite dissolution.
A similarly well-defined sulfate/methane interface coincides with the dissolved sulfide peak at ~40 mbsf. Dissolved methane concentrations slowly rise from 7 µM at 1 mbsf to 55 µM at 35 mbsf. From 40 to 56 mbsf, methane concentrations then rapidly rise to 2 x 103 µM at 56 mbsf and hover in the range of 103 µM for the remainder of the drilled sediment column. The disappearance of almost all methane at the depth of sulfate depletion indicates that most of the methane diffusing upward through this sediment column is ultimately destroyed by anaerobic methanotrophy. The presence of methane at a low concentrations throughout the overlying sediment column indicates, as at open-ocean Sites 1225 and 1226, that methane can be maintained at a background level of several micromolar in subseafloor sediments, despite the potential for methane oxidation by sulfate reduction.
Like methane, ethane and propane are detected throughout most of the sediment column. Ethane is present throughout the sediment column below ~1 mbsf, and propane is present throughout the column below ~11 mbsf. Concentrations of ethane decline sharply at the 40-mbsf top of the anaerobic methanotrophy zone (from 2 to 0.7 µM). Concentrations of propane decline more gradually (from 3 to 0 µM) in parallel with methane across the same interval. These distributions demonstrate that ethane and propane are biologically consumed in the anaerobic methanotrophy zone at this site. Concentrations of all three hydrocarbon species exhibit small distinct peaks in the upper part of the sulfate-rich zone. These small peak occurrences demonstrate that methane, ethane, and propane are all biologically produced in sulfate-rich sediments at this site. Methanogenesis occurs at 1 mbsf, whereas ethanogenesis and propanogenesis occur at ~10 mbsf. Most of the methane, ethane, and propane produced in these sulfate-rich sediments are consumed within a few meters (~5 mbsf for methane and 15-25 mbsf for ethane and propane). Trace concentrations of ethane (10-1 µM) persist throughout the sulfate-rich sediments at this site. This persistence indicates that ethane can be maintained at a very low background level in sulfate-rich sediments, despite its potential for oxidation by sulfate reduction.
In most incubation samples from Site 1227, hydrogen concentrations are between 0.2 and 0.5 nM. These concentrations closely resemble those observed at open-ocean Site 1226. Samples from the first few meters of the sediment column exhibit significantly higher hydrogen concentrations (0.9-2.4 nM). These concentrations are consistent with observation in shallower nearshore sediments (Hoehler et al., 1998). However, similar concentrations in samples from 93 and 113 mbsf are a factor of five to ten lower than those observed in nearshore methanogenic sediments. As for Site 1226, further investigation will be needed to determine whether or not these results indicate that the Site 1227 methanogenic and sulfate-reducing communities utilize hydrogen at free-energy yields lower than the generally accepted theoretical limit for actively growing cells.
The VFAs, formate and acetate, are important intermediates in the anaerobic pathways of organic matter degradation and were analyzed throughout the sediment column. Acetate concentrations range between 0 and ~10 µM and generally increase from the surface sediment down to the base of the drilled sediment column (~150 mbsf). Formate concentrations vary considerably throughout the sediment column (between 0 and ~5 µM) but exhibit no mean trend over the sampled sediment column. The average acetate and formate concentrations of this site are an order of magnitude higher than concentrations in sediments of the equatorial Pacific sites and are similar to concentrations found in very active coastal marine sediments. These results suggest that relative substrate concentrations of different sites may be related to the activity levels of the principal prokaryotic processes, although the absolute process rates are orders of magnitude lower in the open-ocean sediments than in the coastal sediments.
Concentrations of dissolved manganese and iron are, respectively, 0-6 and 0-30 µM at Site 1227. The peak manganese concentration from Site 1227 (6 µM) is a factor of 27 lower than that of equatorial Pacific Site 1225 and a factor of 7 lower than that of equatorial Pacific Site 1226. The peak iron concentration from Site 1227 (30 µM) is a factor of 1.3 greater than that at Site 1225 and a factor of 1.5 lower than that at Site 1226. There are at least two possible explanations why the dissolved iron and manganese concentrations are low at Site 1227 relative to the open-ocean sites. Either the ferrimagnetic material at this ocean-margin site is not an effective source of bioavailable manganese and iron oxides, or dissolved manganese and iron are scavenged and precipitated much more quickly at this site. Stratigraphic relationships between magnetic susceptibility and dissolved sulfide concentrations suggest that these dissolved metals are scavenged by sulfide precipitation at Site 1227. A relatively steep decline in sulfide concentrations from 40 to 75 mbsf is associated with the prominent magnetic susceptibility peak from 40 to 50 mbsf. The ultimate sink for sulfide diffusing deeper into the column is associated with the other most prominent magnetic susceptibility peak at this site (which begins at ~140 mbsf).
A pronounced peak in the values of almost every physical property measured at this site spans the interval from 40 to 50 mbsf. These physical properties include magnetic susceptibility, GRA bulk density, grain density, P-wave velocity, NGR, thermal conductivity, and axial formation factor. Smaller peaks in the values of most of these properties are present in the uppermost 20 m of the sediment column. The bulk porosity profile mirrors the variability in other physical properties at this site; its downhole record is nearly the exact inverse of the bulk density and grain density records. These variations in physical properties result from variations in the bulk lithology of the sediment column. The porosity lows and high values in other physical properties are present in sandier intervals of the sediment column.
The 40- to 50-mbsf interval is composed of sandy silt, rich in glauconite, dolomite, quartz, feldspar, pyrite, and shell fragments. It grades upward into dolomite-bearing clayey silt, rich in diatoms and nannofossils. It directly overlies clay- and nannofossil-bearing diatom ooze. Traces of bioturbation are much more abundant in the 40- to 50-mbsf interval than in the overlying and underlying sediments. The primary front of active anaerobic methanotrophy occurs at the top of this 40- to 50-mbsf sandy interval. The successive fronts of barite precipitation and barite dissolution are present in the same interval. Peak concentrations of dissolved iron, manganese, silica, and phosphate are also present in this interval. Secondary peaks in the dissolved concentrations of iron, manganese, silica, and phosphate are present between 0 and 20 mbsf and are similarly associated with relatively coarse-grained sediments. These relationships suggest that several principal activities of the subsurface biosphere (including anaerobic methanotrophy, iron and manganese reduction, ethanotrophy, and propanotrophy) are pinned in a narrow stratigraphic interval by physical properties and sediment composition at this site. Its mineral composition and its traces of relatively intensive bioturbation indicate that the physical and compositional properties of this interval are primarily determined by the nature of the sediment when it was first deposited on the seafloor. However, to some extent, these properties may have been modified by the postdepositional microbial activities that still occur in them today. Density and porosity can be affected by biologically mediated precipitation and dissolution of authigenic minerals, such as barite, dolomite, and apatite. Magnetic susceptibility may be diminished by biologically mediated dissolution of solid-phase iron oxides and subsequent iron reduction. To a much lesser extent, magnetic susceptibility may also be enhanced by massive biologically induced precipitation of reduced iron and manganese. More detailed determination of the extent to which physical and compositional properties control the microbial activities at this site and the extent to which those activities control the physical and compositional properties will require further investigation.
Preliminary cell counts of eight samples from Site 1227 suggest that sedimentary cell concentrations at most sediment depths are slightly higher at this ocean-margin site than at equatorial Pacific Site 1225. Based on the same few data, at most sediment depths, cell concentrations from Site 1227 may be roughly equivalent to those of open-ocean Site 1226. This data set will be expanded by postcruise analyses.
Experiments on major microbial processes and on enumeration of viable prokaryotes were initiated at selected depths ranging from near the mudline to the bottom of the drilled sediment column. The studied processes include methane and acetate formation and consumption, sulfate reduction, hydrogen oxidation, and rates of cell growth. The cultivation experiments include selective growth conditions for a wide range of autotrophic and heterotrophic prokaryotes ranging from psychrophilic to thermophilic. Specific sampling was targeted to the sulfate/barium interface to study the possible attack of sulfate-reducing bacteria and methane-oxidizing archea on sulfate bound in barite.
One Adara and two DVTP deployments combined with the Leg 112 data define a linear gradient of 49°C/km, with a sediment/water interface temperature of 8.6°C and an estimated temperature at 160 mbsf of 16.4°C. Throughout the sediment column, temperatures are in the low mesophilic range.
Trials were undertaken of three experimental tools at this hole: the pressure coring sampler (PCS), the DVTP-P, the Fugro percussion corer (FPC), and the APC-Methane (APC-M) tool.
Background and Objectives. Site 1228 was one of three Leg 201 sites selected for drilling on the continental shelf of Peru. These shelf sites were collectively selected to provide records of microbial activities, communities, and geochemical consequences in organic-rich ocean-margin sediments.
The principal objectives at this site were
Site 1228 is in the immediate vicinity of Leg 112 Site 680. As described in "Principal Results" in "Site 1227" geochemical studies of Leg 112 sites show that brine is present below the seafloor in the Trujillo and Salaverry Basins (Suess, von Huene, et al., 1988). Interestingly, at Site 680 the deep brine source of sulfate prevents the interstitial water concentration of sulfate from becoming depleted at any depth. Site 1228 therefore provides an opportunity to study how the introduction of sulfate-bearing brine affects subseafloor life in organic-rich, sulfate-rich sediments. Consequently, it provides an excellent standard of comparison for Sites 1227 and 1229, which are, respectively, affected by the intrusion of sulfate-free brine and sulfate-rich brine into organic-rich, sulfate-depleted sediments.
Site 1228 is located at 252 m water depth on the outer shelf edge in the middle of the modern oxygen minimum zone of the Peruvian high-productivity upwelling system. At this depth on the Peru shelf, an oxidized sediment zone is practically absent at the sediment/water interface and sulfate reduction is the predominant mineralization process to the very surface (Rowe and Howarth, 1985; Fossing, 1990; Parkes et al., 1993). The organic content is high at Site 680 (3%-10% TOC), and sulfate reduction rates are still detectable with radiolabeled sulfate in samples taken from as deep as 80 mbsf (Parkes et al., 1990).
The lithologic and physical properties at Site 680 change strongly through the 200-m-deep interval drilled during Leg 112 (Shipboard Scientific Party, 1988a). The sediment mainly consists of diatom mud in the upper 50 m of the Pleistocene deposit. Below 50 mbsf, the terrestrial component of the mud is higher but the sediment is primarily biogenic. The lower part of the sediment column consists of a coarse-grained phosphate and feldspar gravel interpreted as drilling artifacts overlying coarse-grained sand cemented by dolomite (Shipboard Scientific Party, 1988a). Dolomite is the primary authigenic phase, but calcite and apatite are also common.
Shipboard chemical analyses from Leg 112 indicate that concentrations of methane at Site 680 are in the range of 10-100 µL/L (0.4-4 µM) in the upper 100 m of the sediment column. Methane was not analyzed at greater depths at Site 680. Concentrations of dissolved sulfate decline from a near-seawater value to a minimum of 6 mM over the uppermost ~50 mbsf and then rise toward higher values in the underlying sediment as a result of diffusion from the underlying sulfate-rich brine (Shipboard Scientific Party, 1988a). A peak sulfide concentration is present between 20 and 40 mbsf (Mossman et al., 1990). Sulfide concentrations were not measured in deeper portions of the underlying brine-affected interval.
Chloride concentrations increase steadily to the base of the hole, and ammonium steadily increases to at least 80 mbsf. Alkalinity exhibits a maximum value at 20 mbsf. Concentrations of calcium and magnesium exhibit minimum values at 5 and 20 mbsf, respectively, and then increase steadily to the base of the hole. The magnesium/calcium ratio peaks at ~5 mbsf and also steadily declines to the base of the hole (Shipboard Scientific Party, 1988a; Kastner et al., 1990).
These downhole profiles of dissolved chemical concentrations are inferred to result from relatively high levels of biological activity throughout the sediment column, coupled with diffusive exchange with the overlying ocean water and with a sulfate-bearing brine introduced at depth. Prokaryotic cell counts and activities were studied to a depth of 9.1 mbsf at Site 680. Nearly 109 cells/mL were present in all samples analyzed. In MPN cultivation studies, 101 to 105 cells/mL were found to be viable (Cragg et al., 1990; Parkes et al., 1990). The subsurface extent of key electron donors (hydrogen, acetate, and formate) and electron acceptors with standard free-energy yields greater than that of sulfate (oxygen, nitrate, manganese oxide, and iron oxides) was not determined for Site 680.
Principal Results. Continuous APC coring from the seafloor to 200 mbsf enabled high-quality sampling for geochemistry and microbiology throughout the drilled sediment column of Site 1228. Because of the overall predominance of sulfate reduction in the highly sulfidic sediment and the presence of sulfate throughout the sediment column, there were no distinct chemical interfaces to target in the sampling scheme for Site 1228. Concentrations of chloride range linearly from a typical seawater concentration at 0 mbsf to twice the seawater concentration at 200 mbsf. This linear profile demonstrates the long-term stability of brine diffusion and provides a reference for all other interstitial water constituents. Analyzed nonconservative species that are affected by microbial activity in the subsurface included sulfate, DIC, and ammonium. Interstitial water analyses at high depth resolution show unexpected details with implications for both the rates of long-term processes and for more recent changes.
Sulfate reduction in the upper 50 m of the sediment column is not sufficient to deplete sulfate at depth. The overall sulfate distribution shows a steep drop in concentrations over the first few meters below the sediment/water interface, a sigmoidal curve in dissolved sulfate concentrations over the following 10 m, a decrease to 2.5 mM at 38 mbsf, and then a continuous increase to 30 mM at 200 mbsf. The sigmoidal curve of the first 10 m indicates that the near-surface distribution of sulfate reduction and/or transport processes changed strongly in geologically recent time and that diffusion through the sediment column has not yet fully adjusted to a new steady state. The continuous increase in sulfate concentrations from 40 to 200 mbsf results from upward diffusion of the underlying sulfate-rich brine.
The depth profiles of DIC and ammonium closely match the described sulfate distribution. The overall DIC profile reveals a distinct DIC maximum of 19 mM at 2 mbsf, a decline to 15 mM, a rise to a second, broader maximum of 20 mM at 25 mbsf, and then a gradual downhole decrease to 4 mM. Ammonium similarly increases from ~2000 µM near the sediment surface to a local maximum of 2600 µM at 2 mbsf, declines slightly, and then increases gradually to 5000 µM downhole. Comparison to Site 680 biochronostratigraphic data (Shipboard Scientific Party, 1988a) suggests that the sediment that contains the DIC and ammonium maxima may have been deposited a few tens of thousands of years ago. These near-surface interstitial water anomalies indicate that steady-state diffusion of biologically active chemicals past the upper sediment column was disrupted by late Pleistocene environmental change and has not yet fully recovered. The exact nature of these changes will be analyzed when a more complete data set becomes available.
Concentrations of manganese and iron in the interstitial water are extremely low (<0.1 µM) down to ~60-80 mbsf. Below this depth, they increase gradually to ~10 µM (manganese) and 50 µM (iron) at 200 mbsf. The source of these dissolved metals at depth may be either diffusion from below or in situ manganese or iron reduction in the lower sediment column.
In contrast to most other ocean-margin sites, including Site 1227, a sulfate/methane interface is absent from the sediment of Site 1228. Methane concentrations remain low throughout the 200-m sediment column, reaching a maximum of only 8 µM. Yet, the distribution of methane clearly reflects the sulfate distribution, with a maximum coinciding with the sulfate minimum and a general inverse correlation between sulfate and methane concentrations throughout the sediment column. These results indicate that even at a concentration above 9% of its seawater level (minimum = 2.5 mM; seawater = 28.9 mM), sulfate regulates the ability of methane-oxidizing consortia to take up methane and maintain low background concentrations. In this respect, Site 1228 provides a unique opportunity to analyze the energetics of anaerobic oxidation of methane and to test current theories of the limiting parameters for this key microbial process.
Acetate and formate are important fermentation products as well as substrates for sulfate-reducing bacteria. Their concentrations in this organic-rich shelf sediment are tenfold higher than those in deep-sea sediments of the tropical Pacific (Sites 1225 and 1226) but only about one-half their concentrations at Peru shelf Site 1227. The Site 1228 data show considerable scatter with depth. Acetate concentrations fall mostly in the range of 1-4 µM and formate concentrations in the range of 0.5-3 µM. The higher concentrations of both fatty acids are present below 100 mbsf. These concentrations are regulated by uptake mechanisms that are not yet fully understood.
Interestingly, the depth of the distinct sulfate minimum at ~40 mbsf is present in an interval of strong lithologic and physical change. At this depth, the sediment shifts from a diatomaceous silt of predominantly hemipelagic origin to older quartz- and feldspar-bearing silt with a more abundant terrestrial component. At 43 mbsf, there is a distinct minimum in porosity and maxima in density, thermal conductivity, and magnetic susceptibility. It is intriguing to speculate that such a physical boundary may temporarily lock the position of biogeochemical zonations in the sediment column.
The temperature gradient in the Site 1228 sediment column was defined from two discrete temperature measurements taken with the DVTP. The results were combined with Leg 112 data to define a linear temperature gradient of 34°C/km and a heat flow of 32 mW/m2. This heat flow estimate is lower than the 46 mW/m2 estimated for Site 680 by the Leg 112 Shipboard Scientific Party (1988a) and confines the previous broad estimate of 20-70 mW/m2 for this site (Yamano and Uyeda, 1990). The temperature increases down through the sediment column from an estimated annual mean of 12.5°C at the seafloor to an extrapolated 19.3°C at 200 mbsf. These temperatures are all within the low mesophilic range for prokaryotes.
Samples were taken for total counts, viable (MPN) counts, and isolations of prokaryotes from selected depths throughout the sediment column. Because of the short transfer time between Sites 1227, 1228, and 1229, the AODCs of total prokaryotic cell numbers at Site 1228 will be conducted postcruise. A large number of MPN samples and isolation incubations target a broad physiological spectrum of heterotrophic and autotrophic prokaryotes that utilize diverse electron acceptors and donors in their energy metabolism. The selective influence of increased salinity and brine composition is also targeted in some incubations. The expected slow growth of deep subsurface prokaryotes will require long postcruise incubation of samples before definite results are obtained from these experiments. This is also the case for the many experiments on microbial processes measured by radiotracer techniques on samples taken from throughout the entire sediment column.
Because the absence of prokaryotic cell contamination from drilling and sampling operations is critical for the isolation of indigenous prokaryotes and measurement of their activities, a PFT tracer was continuously added to the drill water. Tracer samples were taken on the catwalk or in the laboratory from all core sections and subsamples used for microbiology. It was demonstrated that PFT concentrations are typically higher at the periphery than at the sampled center of whole-round core segments. The microbiology subsamples had PFT concentrations below or near the detection limit. This limit corresponds to the potential introduction of 0.01 µL seawater/g sediment. Such seawater introduction could maximally introduce 5 cells/g, based on the mean cell density in seawater (5 x 10-8 cells/L). An additional contamination test uses fluorescent microbeads dispersed on impact at the head of the core barrel. At Site 1228, this test consistently indicates that contamination is unlikely. This method releases 2 x 1011 prokaryote-sized beads at the most sensitive position during drilling, and the tests on microbiological samples showed no beads or, at most, one bead in the >60 microscopic fields of view routinely scanned. The extensive contamination tests applied at this site thus confirm the high quality of microbiology samples that can be taken by careful techniques from APC cores without visible disturbance.
Background and Objectives. Site 1229 was one of three Leg 201 sites selected for drilling on the continental shelf of Peru. These shelf sites were collectively selected to provide records of microbial activities, communities, and geochemical consequences in organic-rich ocean-margin sediments.
The principal objectives at this site were
Site 1229 is located on the Peru shelf in 150.5 m water depth. It is in the immediate vicinity of Leg 112 Site 681. As described in "Principal Results" in "Site 1227" geochemical studies of Leg 112 sites show that brine is present below the seafloor in the Trujillo and Salaverry Basins (Suess, von Huene, et al., 1988). Site 1229 provides an opportunity to study how the occurrence of sulfate-bearing brine affects subseafloor life in organic-rich, sulfate-depleted, methane-rich sediments. Consequently, it provides an excellent standard of comparison for Sites 1227 and 1228, which are, respectively, affected by the intrusion of sulfate-free brine into organic-rich, sulfate-depleted sediments and the intrusion of sulfate-rich brine into sediments with sulfate-bearing interstitial waters.
Shipboard chemical analyses from Leg 112 indicate that concentrations of methane at Site 681 increase from 102 to 105 µL/L in the first 40 m of the sediment column and decline from 105 to 102 µL/L between 73 and 100 mbsf (Shipboard Scientific Party, 1988b). In contrast, the concentrations of dissolved sulfate decline to 0 mM over the first ~30 mbsf, remain at or near 0 mM until 75 mbsf, and then increase steadily with greater depths (Shipboard Scientific Party, 1988b). This downhole pattern of sulfate concentrations indicates active sulfate reduction at depths above 30 mbsf and at depths below ~75-100 mbsf. The downhole pattern of methane concentrations indicates that methane is created at depths of 60-70 mbsf and diffuses to the overlying and underlying zones of active sulfate reduction, where both sulfate and methane are destroyed.
Chloride concentrations increase steadily to the base of the hole. Ammonium concentrations decline slightly from the sediment/water interface to 12 mbsf, increase from 12 to 80 mbsf, and then begin to decline again. Alkalinity also declines from the sediment/water interface to 12 mbsf, increases to a subsurface maximum at 32 mbsf, and then declines again with depth. Calcium and magnesium concentrations exhibit minimum values at ~30 mbsf and then increase steadily to the base of the hole. The magnesium/calcium ratio exhibits a broad peak from ~0 to 40 mbsf and then steadily declines to the base of the hole (Shipboard Scientific Party, 1988b).
These downhole profiles of dissolved chemical concentrations are collectively inferred to result from high levels of biological activity and biologically driven solid-phase alteration throughout the sediment column, coupled with diffusive exchange with the overlying ocean and with a sulfate-bearing brine introduced at depth. AODCs show that prokaryotic cells are present in samples taken from as deep as 80 mbsf at Site 681 (Cragg et al., 1990). Viable prokaryotes were found and potential activity rates were identified in the same samples (Cragg et al., 1990). The subsurface extent of key electron donors (hydrogen, acetate, and formate) and electron acceptors with standard free-energy yields greater than that of sulfate (oxygen, nitrate, manganese oxide, and iron oxides) was not determined for Site 681.
Principal Results. An important objective for Site 1229 is to identify and quantify zones of microbial activity based on reactive interstitial water species. Toward this end, we established a highly resolved chemical record throughout the drilled sediment column. Profiles of conservative ions provide evidence of diffusive mixing between seawater diffusing downward from the sediment/water interface and a brine diffusing upward from older sediments. For example, concentrations of dissolved chloride increase linearly from 559 mM at the sediment/water interface to 1208 mM at the base of the drilled sediment column (186 mbsf). Peak concentrations of biologically affected chemical species, such as ammonium (5800 µM) and dissolved inorganic carbon (22 mM), indicate that rates of subseafloor microbial activity are much higher at this ocean-margin site than at open-ocean Sites 1225 and 1226. These peak concentrations also suggest that the subseafloor microbial activity at Site 1229 is slightly greater than that at Site 1228 (which lies just seaward of Site 1229) and perhaps is slightly less than that at Site 1227 (which is situated 310 km to the north on the Peru shelf).
As at Site 1228, the concentration profiles of several biologically affected chemical species exhibit a pronounced anomaly just below the seafloor (at 2-3 mbsf). This anomaly at Site 1229 consists of a brief positive excursion in alkalinity, DIC, ammonium, and sulfide, with a co-occurring negative excursion in dissolved sulfate. The same anomaly is also apparent in the ammonium and alkalinity profiles of Site 681 (Shipboard Scientific Party, 1988b). As described in "Principal Results" in "Site 1228" this near-surface interstitial water anomaly indicates that the steady-state diffusion of biologically active chemicals past the upper sediment column was disrupted by late Pleistocene environmental change and has not yet fully recovered. There are least three possible general explanations of this anomaly. It may result from ongoing activity in a microbial "hotspot" at this shallow sediment depth, it may be a chemical relic of past microbial activity (now relaxing back to a diffusional steady state), or it may be a result of the recent establishment of an oxygen minimum at this water depth, causing the extinction of a bioirrigating benthos and a stimulation of sulfate reduction in the uppermost 2 m of sediment.
The most striking biogeochemical feature of this site is the reversal of the biogeochemical zonation at depth. This reversal is immediately apparent in the dissolved sulfate profile. The sulfate concentrations decline from a seawater value of 29 mM at the sediment surface to 0 mM at ~35 mbsf. They remain at 0 mM from 35 to 88 mbsf and then steadily rise from 0 to 38 mM at 186 mbsf. The sulfate that sustains microbial reduction over the uppermost 35 mbsf of the sediment column ultimately diffuses downward from the overlying ocean. The sulfate that sustains microbial reduction below 88 mbsf is inferred to diffuse upward from the underlying brine. Both intervals of sulfate reduction are marked by local maxima in the concentrations of dissolved sulfide, with a broad peak from ~20 to 40 mbsf and a sharper peak at ~90 mbsf.
The sulfate profile is mirrored by the dissolved methane profile. Dissolved methane concentrations are <100 µM from 0 to 20 mbsf, hold steady at a few hundred micromolar from 20 to 35 mbsf, and then rise to values of ~2000 µM (exceeding 1 bar partial pressure) between 65 and 75 mbsf. Methane then steadily declines to <100 µM at 93 mbsf and remains in the range of 100 µM or less to the base of the sampled sediment column. As at Site 1227, the disappearance of almost all methane at the depths of sulfate depletion indicates that most of the methane in this sediment column is ultimately destroyed by anaerobic methanotrophy. As observed at all previously drilled Leg 201 sites, the Site 1229 methane and sulfate profiles indicate that methane can be maintained in subseafloor sediments at background concentrations that are inversely related to the co-occurring dissolved sulfate concentrations.
The dissolved iron and manganese concentration profiles demonstrate that net reduction of iron and manganese oxides occurs in the methanogenic zone. The principal foci of net manganese and iron reduction are at slightly different depths, with iron reduction peaking at 75-90 mbsf and manganese reduction just above and below that interval. The presence of methanogenesis in iron- and manganese-reducing environments may result from a limited availability of mineral-supplied electron acceptors relative to electron donors. In these organic-rich sediments, electron donors may be supplied to the microbial community faster than mineral dissolution can supply dissolved reducible manganese and iron. Relatively high concentrations of manganese and iron in the lower sulfate zone could be due to either in situ mineral reduction or to diffusion from the underlying brine-rich sediment.
The dissolved barium profile is broadly similar to the methane profile. Dissolved barium concentrations are <2 µM from 0 to 24 mbsf. Concentrations of barium in interstitial water then rapidly rise to 18 µM at 40 mbsf and remain near 19 µM until almost 80 mbsf. They then decline steeply to 2 µM at ~100 mbsf and are <2 µM for the remainder of the drilled sediment column. As at Site 1227, the inverse relationship between sulfate and barium is inferred to be controlled by the solubility product of barite. Upward diffusion of barium past 35 mbsf and downward diffusion of barium past 90 mbsf is suspected to sustain modern barite formation at, respectively, ~24 and 100 mbsf. Similarly, the shoulders of the barium peak at ~40 and 80 mbsf are inferred to mark the principal depths of current barite dissolution at this site.
Prokaryotic cell counts were done at 10-m intervals throughout the upper sediment column and across both sulfate/methane interfaces. These data show that mean sedimentary cell concentrations are several-fold higher at this ocean-margin site than at the Leg 201 open-ocean sites and may be slightly higher than mean concentrations at nearby Site 1227. The most striking features of the shipboard cell counts are the thousandfold increase in cell concentrations in the lower zone of overlapping sulfate and methane concentrations and the tenfold increase in cell concentrations in the upper zone of overlapping concentrations. The maximum cell concentrations observed in the lower sulfate/methane zone are actually an order of magnitude higher than the concentrations observed at the sediment/water interface. Given the coarse spacing of these samples and their positions relative to the chemically defined sulfate/methane overlap zones, the peak cell concentrations observed in the upper sulfate/methane zone may greatly underestimate the peak concentrations in that zone.
Acetate and formate concentrations exhibit strong local maxima of ~6 µM in both of the sulfate/methane interface zones. These maxima are centered at 37 and 90 mbsf. As with the cell counts, these local maxima are higher than the local maxima exhibited by both acetate (~2 µM) and formate (3 µM) at the sediment/water interface. Throughout most of the remaining record at this site, concentrations of both species are between 1 and 2 µM. As at Site 1227, the concentrations of both species reach their highest values near the base of the drilled sediment column (~15 µM). These results are intriguing because these volatile acids are important substrates for both sulfate reducers and methanogens. Hydrogen is another important electron donor in anaerobic communities. Almost all hydrogen concentrations measured at this site were <0.5 nM, and most were <0.2 nM. These concentrations resemble those observed at open-ocean Site 1225 and ocean-margin Site 1227. As noted in "Principal Results" in "Site 1225" and "Principal Results" in "Site 1227" these concentrations are much lower than expected from experiments with sulfate-reducing and methanogenic communities of surface sediments. The accurate interpretation of these acetate, formate, and hydrogen concentrations must await postcruise analyses of prokaryotic energetics in subseafloor environments.
The cell concentration data and sulfate and methane gradients demonstrate that the subseafloor prokaryotic population and activity are locally strongly focused at the sulfate/methane overlap zone defined by the upward-diffusing sulfate-bearing brine and the downward-diffusing seawater sulfate. The dissolved barium profile indicates that microbial activity in this zone directly influences sediment chemistry by mediating the precipitation and dissolution of barite. In these effects on subsurface biological activities and biogeochemical cycles, this brine-caused sulfate/methane interface mirrors the effects of the overlying "normal" sulfate/methane interface. Postcruise microbiological studies will be required to demonstrate whether or not the microbial community supported by the brine-induced interface is locally unique or the same as that supported by the overlying interface.
The upper sulfate-rich zone at Site 1229 lies entirely within lithostratigraphic Subunit IA, a stratigraphic interval of primarily hemipelagic sediments (0-40 mbsf). The underlying methane-rich zone is largely limited to lithostratigraphic Subunit IB, which is the upper portion of a longer interval (40-138 mbsf) of mixed terrigenous and hemipelagic sediments. The AOM zones that separate the upper and lower sulfate-rich zones from the intervening methane-rich zone are associated with brief sedimentary intervals characterized by high grain density, high NGR, high resistivity, and low porosity. These brief low-porosity intervals are unusually rich in terrigenous sediment and are interpreted to have been deposited during the two most recent lowstands of four onlap/offlap cycles that define the 40- to 138-mbsf interval.
In short, as at Site 1228, the upper sulfate-reducing interval at Site 1229 is composed of predominantly hemipelagic sediments, the strongly methanogenic zone is rich in terrigenous sediment relative to the overlying sulfate-reducing zone, and the intervening AOM zone is present just above an interval of low-porosity, high-density lowstand sediments. The lower AOM zone at Site 1229 is present within a similar interval of high-density, low-porosity lowstand sediments. The lithologic association of AOM zones with high-density, low-porosity lowstand sediments at Sites 1229, 1228, and 1227 provides intriguing evidence that, on the Peru shelf, the position of AOM zones is currently pinned within the sediment column by lithologic properties and, by extension, depositional history.
As at Site 1227, stratigraphic patterns of magnetic susceptibility and dissolved manganese, iron, and sulfide concentrations indicate similar control of other microbial processes by depositional history at Site 1229. Magnetic susceptibility is generally much higher in the methanogenic zone and in the lower sulfate-reducing zone than in the overlying sulfate-reducing zone. This circumstance suggests that mineral sources of reducible iron and manganese are more abundant in the terrigenous-dominated sediments of the lower sulfate-reducing zone and the mixed terrigenous and hemipelagic sediments of the methanogenic zone than in the mostly hemipelagic sediments of the upper sulfate-reducing zone. The relatively high magnetic susceptibility of the intervals with more strongly terrigenous sediments is consistent with our finding that dissolved manganese and iron concentrations are generally higher in the lower methanogenic zone and the underlying sulfate-reducing zone than in the upper sulfate-reducing zone. The presence of higher manganese and iron concentrations and lower sulfide concentrations in these relatively high-susceptibility intervals in turn provides strong evidence that the current rates and stratigraphic foci of iron reduction, manganese reduction, and sulfide precipitation depend strongly on depositional history.
Experiments on major microbial processes and on enumeration of viable prokaryotes were initiated at selected depths ranging from near the mudline to the bottom of the drilled sediment column. The studied processes include methane and acetate formation and consumption, sulfate reduction, hydrogen oxidation, and rates of cell growth. The cultivation experiments include selective growth conditions for a wide range of autotrophic and heterotrophic microorganisms ranging from psychrophilic to thermophilic. Detailed microbiological sampling targeted the top of the sediment column and both the upper and lower sulfate/methane overlap zones.
The results from one DVTP deployment were combined with temperature data from Site 681 to define a linear gradient of 35.5°C/km for this site. The mean sediment/water interface temperature defined by this gradient is 13.4°C. The temperature defined for the base of the drilled sediment column (193 mbsf) is 20.2°C. Throughout this interval (0-193 mbsf), temperatures are in the low mesophilic range.
Trials were undertaken of four experimental tools at this hole: the PCS, the DVTP-P, the APC-M tool, and the FPC.
Background and Objectives. Site 1230 was the single hydrate-bearing site selected for drilling during Leg 201. The principal objectives at this site were
Site 1230 is located on the lower slope of the Peru Trench in 5086 m water depth. Sediments of this area are part of the accretionary wedge just landward of the Peru Trench (Suess, von Huene, et al., 1988). The lithologies, sediment age, and many geochemical and geophysical characteristics of the target site were well characterized by Leg 112 studies of nearby Site 685 (Shipboard Scientific Party, 1988d). The upper 200 m of Pleistocene to Holocene sediment is a clay-rich diatomaceous mud, partly accreted by downslope transport from the shelf. At ~ 200 mbsf, a stratigraphic hiatus of ~4.5 m.y. separates the slope deposit from upper Miocene diatom ooze (Shipboard Scientific Party, 1988d). Authigenic carbonates and phosphates are sparse, whereas pyrite framboids are abundant throughout the section (Shipboard Scientific Party, 1988d). Calculated sedimentation rates are high; they average 250 m/m.y. for the Miocene sequence and 100 m/m.y. for the Pleistocene section (Shipboard Scientific Party, 1988d). These high rates are consistent with sedimentation in a lower-slope basin or trench axis.
The surface waters over Site 1230 are part of the Peru upwelling system and are biologically highly productive. The organic carbon content of the sediment is high at Site 685 (Shipboard Science Party, 1988d). Methane concentrations were observed to rise above 1 bar by 11.6 mbsf and remain in the range of 104-105 µL/L throughout the cored sediment column down to 432 mbsf (Kvenvolden et al., 1990). Concentrations of ethane and butane generally increase downhole from 1 to 100 µL/L, and the methane/ethane ratio decreases from 105 to 103. The Leg 112 Scientific Party found visual evidence of methane hydrate at 99 and 164 mbsf in the form of small pieces of dark gray hydrate (Shipboard Scientific Party, 1988d; Kvenvolden and Kastner, 1990). The samples looked like rounded pieces of mudstone but felt cold and showed bubbling foam. Based on this information, Site 1230 provides an excellent opportunity for assessing the nature of microbial communities and their activities in hydrate-bearing sediments rich in organic material and under high hydrostatic pressure.
Concentrations of dissolved sulfate declined to 0 mM between the first and second core analyzed at Site 685 (between 3 and 18.1 mbsf) (Shipboard Scientific Party, 1988d). Chloride concentrations range between 525 and 555 mM. The maximum concentration is associated with the most shallow sulfate-free sample (18.1 mbsf) and was suggested by the Leg 112 Shipboard Scientific Party to lie just above hydrate at the top of the hydrate stability field. Salinity, alkalinity, dissolved ammonium, phosphate, and magnesium concentrations rise to maximum values in the interval of 107-134 mbsf, decline sharply between 165 and 235 mbsf, and then decrease gradually to the base of the hole at ~450 mbsf. The maxima in alkalinity (156 mM), ammonium (31,760 µM), and phosphate (826 µM) were the highest then known from deep-ocean drilling (Shipboard Scientific Party, 1988d). Downhole variation in chloride and calcium concentrations is generally opposite to the variation in these other chemical species. The pH drops below 7 at 133 mbsf and remains below 7 to the base of the hole (Shipboard Scientific Party, 1988d).
These patterns of interstitial water chemistry are inferred to result from high levels of biological activity throughout the sediment column, coupled with hydrate formation and diffusive exchange with the overlying ocean. The subsurface extent of key electron donors (hydrogen, acetate, and formate) and electron acceptors with standard free-energy yields greater than that of sulfate (oxygen, nitrate, manganese oxide, and iron oxides) was not determined for Site 685.
Principal Results. The biogeochemical zonation of Site 1230 is more typical of an upper-slope sediment than a typical deep-sea sediment; its uppermost sediment contains a narrow suboxic zone, and sulfate depletion occurs at <9 mbsf. Oxygen and nitrate are not detectable at the top of the mudline core. Dissolved manganese is present in the uppermost 0.5 m of sediment but is near the detection limit (<1 µM) throughout the remaining sediment column. Dissolved iron is likewise low (mostly 1-3 µM) in the upper 25 m of the sediment. Below the narrow suboxic zone, sulfate reduction is the dominant mineralization process down to the bottom of the sulfate zone at 8-9 mbsf. The sulfate gradient is nearly linear and indicates that most of the net sulfate reduction takes place at the sulfate/methane interface (Iversen and Jørgensen, 1985; Niewöhner et al., 1998; Borowski et al., 1996, 2000).
Methane builds up steeply beginning at the sulfate boundary, and it reaches >1 bar partial pressure by 11 mbsf. Below that depth, methane concentrations in recovered cores fluctuate around a few millimolar, which is the usual pattern in supersaturated cores with gas escape upon depressurization. At Site 1230, however, nine successful deployments of the PCS at depths ranging from 22 to 277 mbsf allowed the methane concentration profile from the entire sediment column to be accurately determined. The PCS recovered a full 1-m core in most deployments. Its highest internal pressure was 8086 psi in a core recovered from 254.6 mbsf. At 254.6 mbsf, 8086 psi is equivalent to 105% of hydrostatic pressure. The overpressure is caused by dissolving gas hydrate resulting from warming during the wireline trip (Dickens et al., 2000). The total amount of methane retrieved by the PCS reached 400,000 µM methane at 157 mbsf. This greatly exceeds methane solubility at the ambient temperature and hydrostatic pressure but is consistent with the presence of several percent gas hydrate in the sediment pore space.
The presence of gas hydrate was also monitored by rapid IR scanning of the recovered cores. Immediately after retrieval, each core was brought to the catwalk and scanned along the core liner surface with a digital IR camera. Our purpose was to detect the cooling effect caused by rapid gas hydrate dissolution. This approach was successful, as core segments with negative temperature anomalies of about -5°C proved to harbor gas hydrate. Hydrate was recovered from ~82 and ~148 mbsf as small pieces mixed with sediment. The recovered hydrate probably represented only a small fraction of the in situ hydrate because of rapid dissolution and loss in the expanding cores. Samples from four additional horizons (123, 142, 150, and 200 mbsf) probably contained disseminated gas hydrate, based on observed fizzing and scanned temperature anomalies as low as -3.2°C. Downhole sonic and resistivity logs suggest broad intervals of possible hydrate presence. Preliminary comparison of inferred hydrate distributions and PCS methane data suggests that the interstitial concentrations of dissolved methane build up to reach the phase boundary of hydrate formation at ~50 mbsf. The dissolved concentrations may remain at this phase boundary at depth, with intervals of hydrate formation determined by the lithology and physical properties of the sediment.
The depth distribution of chloride in the interstitial water also provided evidence of hydrates, which release freshwater by dissolution during the wireline trip of the sediment core. Chloride shows a distinct gradient with a peak at 18 mbsf. This subsurface peak is presumably a remnant of the last glacial salinity excursion. It is accentuated by a drop in chlorinity below 18 mbsf that is probably due to freshening by hydrate dissolution. Within the methane zone, the drop in chlorinity is 10-27 mM and the concentrations show strong depth fluctuations with minimum values that appear to coincide with depths of hydrate occurrences (e.g., at 82 mbsf).
Ethane and propane are present at 1-2 ppm concentrations throughout the methane-rich zone down to ~140 mbsf. Their concentrations increase three- to fivefold over the next 70 m. Their distribution profiles suggest that ethane and propane are products of organic carbon degradation in the methanogenic zone.
Interstitial water analyses at Site 1230 provide clear evidence of very high microbial activity with extreme accumulations of products from organic degradation processes. Alkalinity and DIC increase steeply with depth from near seawater values at the sediment/water interface to a broad maxima of 155 mM at 100-150 mbsf, deep in the methanogenic zone. These concentrations are among the highest ever measured in marine sediments. Below this maximum, the concentrations drop again with depth. Ammonium likewise builds up extreme concentrations of 35,000 to 40,000 µM from 100 to 150 mbsf.
Below the interface of counter-diffusing sulfate and methane, there is a second diffusive interface between hydrogen sulfide and iron at 25 mbsf. The hydrogen sulfide produced from sulfate reduction reaches a peak concentration of 9.4 mM at the bottom of the sulfate zone. From there it decreases steeply both upward and downward, reaching zero near the sediment/water interface and at 25 mbsf. Iron is abundant (5-57 µM) in the interstitial water of the methane zone from 200 to 25 mbsf, where it meets the hydrogen sulfide and is inferred to precipitate as ferrous sulfide and pyrite.
A diffusive interface between sulfate and barium is encountered at 8-9 mbsf. Barium concentrations are only a few micromolar in the sulfate zone but increase steeply below that zone to plateau at 400 µM between 50 and 150 mbsf. At 250 mbsf the barium concentration approaches 1000 µM, which may be the highest interstitial water concentration of barium ever recorded in deep-sea sediments. The narrow depth interval of coexisting barium and sulfate appears to be a zone of barite precipitation. We infer their concentrations to be determined by the solubility product of barite in that zone. Consequently, the shallow sulfate zone is an effective barrier against upward diffusion of dissolved barium. Barium fronts associated with the sulfate boundary have also been observed in sediments of the Gulf of California and the South Atlantic Ocean (Brumsack, 1986; Kasten et al., 2001). Based on data from Leg 112, von Breymann et al. (1990) concluded that the deepest sites have the highest dissolved and solid-phase barium concentrations because detritus sedimenting through a deepwater column scavenges barium from seawater and enriches the sediment in barium.
Acetate and formate are generated as fermentation products and are used as substrates by sulfate-reducing or methanogenic prokaryotes. These VFAs are present at much higher concentrations at Site 1230 than at any other site studied during Leg 201. The acetate level is 5-20 µM in the sulfate reduction zone and reaches 230 µM in the methane zone at 145 mbsf. This acetate concentration is fivefold higher than at the most active sites on the Peru shelf and is even 10- to 100-fold higher than at the other deep-sea sites. Formate remains mostly at 5-10 µM throughout the sediment column. Hydrogen concentrations are low, in the 0.1- to 1.5-nM range.
The interstitial water at Site 1230 has a distinct yellow color that is not present at any other Leg 201 site. We presume this color is probably due to dissolved organic matter. The intensity of the color, which was measured spectrophotometrically, increases steeply from zero at the sediment/water interface to a broad maximum between 25 and 150 mbsf. Below that depth, it drops again to reach 15%-20% of the maximum value at 250 mbsf.
Prokaryotic cell concentrations in the organic-rich Pleistocene to Holocene sediments are near the average of previously studied subseafloor sediments in the upper 60 m of the sediment column. They are about threefold above average in the next 150 m. However, in the older accretionary wedge sediments below 216 mbsf, the cell density abruptly drops fourfold, from 7.9 x 106 to 1.9 x 106 cells/cm3. This shows that the concentration of subseafloor prokaryotic cells at Site 1230 is closely related to sediment age rather than sediment depth. The factor that ultimately regulates cell concentrations may be the availability of energy substrates for prokaryotic metabolism.
Samples were taken at regular depth intervals through the entire sediment column for DNA and FISH-SIMS analysis, measurements of sulfate reduction rates, hydrogen turnover, methanogenesis rates, acetate turnover, thymidine incorporation, and prokaryote lipid biomarkers. Samples for cultivations and viable counts (MPN) target specific depths and geochemical zones, including the sulfate/methane interface and the hydrate-rich methane zone. Contamination tests with PFT and fluorescent beads show that the potential seawater contamination of Site 1230 microbiological samples is low or undetectable. The only case of detectable bead contamination in a slurry used for Site 1230 microbial cultivations is based on two beads counted in 100 microscopic fields of view scanned. By the experience accumulated during this leg, our confidence has strengthened that, with rigorous contamination controls and aseptic sampling techniques, deep subsurface samples can routinely be obtained without the introduction of microorganisms from the surface environment.
Four successful temperature measurements (two Adara tool deployments and two DVTP deployments) over a depth interval of 0-255 mbsf defined a geothermal gradient of 34.3°C/km at Site 1230, with a mudline temperature of 1.7°C and an estimated temperature of 11.2°C at 278 mbsf. The estimated local heat flux is 28 mW/m2. This is similar to the heat flux calculated by Yamano and Uyeda (1990) at Site 685 from wireline logging data over 75-150 mbsf. Based on a downhole measurement of overpressure, upward interstitial water advection of ~1 cm/yr may occur at this site.
![]()