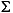 H2S = H2S + HS-) is below the detection limit (0.0002 mM) throughout the entire sediment column.
H2S = H2S + HS-) is below the detection limit (0.0002 mM) throughout the entire sediment column.
At Site 1231, the dissolved inorganic carbon (DIC) profile hovers slightly at or below 3.3 mM for most of the sediment column. It exhibits three slight exceptions to this relative constancy: it slightly increases from 2.8 mM near the sediment/water interface, it exhibits a small peak of ~3.7 mM centered at 55 mbsf, and it declines slightly to ~3.0 mM at the sediment/basement interface. These DIC concentrations are even lower than those at Site 1225 (3.0-4.0 mM). They are much lower than the DIC concentrations observed at the other Leg 201 sites. Dissolved ammonium concentrations are also generally lower at Site 1231 than at the other Leg 201 sites. As at Site 1225, concentrations of DIC, ammonium, and alkalinity peak in the middle of the sediment column and decline toward both the sediment/ocean interface and the sediment/basement interface. The relatively low variability in the concentration profiles of these chemical species suggests that net microbial activity may be lower at Site 1231 than at any other Leg 201 site. The midcolumn peaks in these profiles and their relatively low values near both the sediment/water and sediment/basement interfaces indicate chemical exchange between the sediment and the ocean and between the sediment and the basement.
Dissolved sulfate concentrations are >28 mM at the sediment surface and decrease linearly to 27 mM near the basement. The slight total downhole decrease in sulfate concentrations suggests that Site 1231 is characterized by very low sulfate-reducing activity. Dissolved sulfide (H2S = H2S + HS-) is below the detection limit (0.0002 mM) throughout the entire sediment column.
Electron acceptors with higher standard free-energy yields than sulfate are present throughout most of the sediment column at Site 1231. Dissolved nitrate appears to be present in the uppermost meter and the lowermost 60 m of the sediment column (where it ranges from 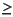 15 µM at 114 mbsf to 2 µM at 77 mbsf). Dissolved oxygen similarly appears to be present in the top 0.6 m below the seafloor as well as the last 3.8 m of sediment above basaltic basement. The diffusion of oxygen and nitrate from the overlying ocean down into the sediment is readily predictable from deep-ocean chemistry. However, the first Leg 201 locations, Sites 1225 and 1226, provided the only previous precedent for upward transport of nitrate (and, at Site 1225, oxygen) into deeply buried sediment from the underlying basaltic crust. As at Site 1225, the introduction of dissolved nitrate high into the sediment column at Site 1231 indicates that nitrate-utilizing microbial activity is present but may proceed at a very low rate in the site's lowermost sediments. Also as at Site 1225, the presence of dissolved oxygen and nitrate in these deepest sediments suggests that microbial activity in the underlying basalt is insufficient to strip even the scarcest preferentially utilized electron acceptors from the water that flows through the basalt.
15 µM at 114 mbsf to 2 µM at 77 mbsf). Dissolved oxygen similarly appears to be present in the top 0.6 m below the seafloor as well as the last 3.8 m of sediment above basaltic basement. The diffusion of oxygen and nitrate from the overlying ocean down into the sediment is readily predictable from deep-ocean chemistry. However, the first Leg 201 locations, Sites 1225 and 1226, provided the only previous precedent for upward transport of nitrate (and, at Site 1225, oxygen) into deeply buried sediment from the underlying basaltic crust. As at Site 1225, the introduction of dissolved nitrate high into the sediment column at Site 1231 indicates that nitrate-utilizing microbial activity is present but may proceed at a very low rate in the site's lowermost sediments. Also as at Site 1225, the presence of dissolved oxygen and nitrate in these deepest sediments suggests that microbial activity in the underlying basalt is insufficient to strip even the scarcest preferentially utilized electron acceptors from the water that flows through the basalt.
Dissolved manganese is present from 1 to 65 mbsf at Site 1231. Concentrations steadily rise from ~17 µM at 1.4 mbsf to a local peak of 78 µM at ~17 mbsf, decline briefly by a few micromolar, and then rise to sustain highest concentrations of 120 µM from 36 to 46 mbsf. Manganese concentrations below this peak steadily decline to essentially 0 µM at 68 mbsf. A relatively broad zone of generally high but variable dissolved iron concentrations (7-36 µM) spans the interval from 1 to 30 mbsf. A very small secondary peak in dissolved iron (5 µM) is centered near 74 mbsf. Two aspects of these broad patterns run counter to the general expectation that manganese reduction occurs at shallower depths than iron reduction in marine sediments. The first aspect is the broad co-occurrence of dissolved iron and manganese from 1 to 30 mbsf. The second aspect is the presence of the maximum dissolved iron concentrations much closer than the maximum dissolved manganese concentrations to the sediment/water interface. It appears likely that rates of manganese reduction in these sediments are limited by the availability of manganese oxides that supply dissolved manganese. Rates of iron reduction may be similarly limited by the presence and solubility of the minerals that supply dissolved iron.
As at other Leg 201 sites, the downhole distribution of microbial manganese and iron reduction at Site 1231 appears to be ultimately determined by lithology and depositional history. The peak intervals of dissolved manganese production are limited to the clays that lie between 11 and 55 mbsf. The maximum manganese concentration (120 µM) is present in the yellow volcanic-rich clay of Subunit IIA (31-44 mbsf). The secondary peak (78 µM) is centered in the green diatom-rich clay of Unit I (11-30 mbsf). Dissolved iron is similarly limited to the clay-rich portions of the upper sediment column. It exhibits a sharp maximum concentration (36 µM) a few meters below the seafloor in the radiolarian and clay-rich diatom ooze of upper Unit I (0-11 mbsf). Most of the dissolved iron at Site 1231 is present in a broad maximum of 26 µM in the green clay of Unit I. Dissolved iron concentrations are consistently 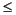 5 µM in the nannofossil oozes that lie between 55 mbsf and the basaltic crust (114 mbsf). Dissolved manganese is consistently <1 µM over the same interval.
5 µM in the nannofossil oozes that lie between 55 mbsf and the basaltic crust (114 mbsf). Dissolved manganese is consistently <1 µM over the same interval.
Although Site 1231 may be the microbially least active of the Leg 201 sites, its sediments still contain methane at concentrations of up to 15 µM. At this site, methane is limited to the upper clay-rich portion of the sediment column between 0 and 42 mbsf. This methane-bearing interval is completely within the interval of high dissolved manganese concentrations. Interestingly, this methane was only detected after prolonged incubation of headspace samples over a couple of days, whereas short 20-min incubation according to the standard Ocean Drilling Program (ODP) safety protocol showed only trace methane concentrations throughout the sediment column. The appearance of methane over time is currently interpreted as a release of sorbed methane. From sediments below 42 mbsf, no release of sorbed methane was observed and the concentrations remained at trace levels of <0.2 µM. The relationship of this sorbed methane to current microbial activity remains unknown.
Acetate concentrations range between 1 and 14 µM at Site 1231. Formate varies between 1 and 19 µM. Concentrations of both fatty acids are lowest in the top 3 m below seafloor (1-2 µM). They are slightly higher (3-6 µM) in the nitrate-reducing zone that spans the last 50 m above basement. Acetate and formate exhibit their highest concentrations (4-14 µM and 9-19 µM, respectively) at intermediate sediment depths (25-75 mbsf and 25-80 mbsf, respectively). These broad patterns suggest that at Site 1231 fatty acid concentrations may be lower in the sedimentary intervals that include electron acceptors with the highest energy yields. Curiously, the acetate and formate concentrations at this site are generally an order of magnitude higher than concentrations in sediments of the equatorial Pacific sites but are similar to those found at the Peru margin sites and in other very active coastal marine sediments. As noted in previous site chapters, accurate understanding of the fatty acid distribution and its microbial relevance will require thorough postcruise analyses of microbial energetics in subseafloor environments.
Hydrogen concentrations are extremely high in the uppermost 35 m of the sediment column, with a peak value of 102 nM at 15 mbsf. This is the highest hydrogen concentration measured at any Leg 201 site. It exceeds the hydrogen concentrations of near-surface iron-reducing sedimentary environments by >100-fold. The zone of high hydrogen coincides with the zone of iron reduction but does not show any direct correlation with distributions of fatty acids or methane. The presence of extremely high hydrogen concentrations at the site with the lowest organic carbon mineralization rates remains unexplained at this point. From 44 mbsf down to the basaltic basement, hydrogen concentrations are, in contrast, very low (0.05-0.22 nM).
Prokaryotic cell counts were conducted on samples from throughout the sediment column at Site 1231. These data show that mean cell concentrations are generally lower at this open-ocean site than at any previously enumerated ocean drilling site. Cell concentrations exhibit a distinct local concentration peak at 10-15 mbsf, the approximate depth of the Unit I zone of iron and manganese reduction.
Experiments on major microbial processes and on enumeration of viable prokaryotes were initiated at selected depths ranging from near the seafloor to the bottom of the drilled sediment column. The studied processes include methane and acetate formation and consumption, sulfate reduction, hydrogen oxidation, and rates of cell growth. The cultivation experiments include selective growth conditions for a wide range of autotrophic and heterotrophic prokaryotes ranging from psychrophilic to thermophilic. Cultivation experiments particularly focused on manganese- and iron-reducing bacteria throughout the column. Studies of sulfate-reducing bacteria in macrofaunal burrows were also initiated. Detailed microbiological sampling targeted sediment depths of particular biogeochemical interest, such as the midcolumn reduced manganese interface and the sediment/basalt interface.
The results from six Adara tool deployments define a temperature profile composed of two distinct intervals: a linear gradient of 90°C/km from 0 to 55 mbsf and a linear gradient of 35°C/km from 55 to 115 mbsf. The sediment/water interface temperature measured by a mudline Adara tool deployment is 1.7°C. The estimated temperature at the base of the drilled sediment column (115 mbsf) is 8.6°C. Throughout the entire drilled interval (0-121 mbsf), temperatures are in the psychrophilic range.
Trials were undertaken of two experimental tools at this site: the Davis-Villinger Temperature-Pressure Probe (DVTP-P) and the catwalk infrared (IR) camera. The single DVTP-P deployment indicated minor overpressure at 108 mbsf.
![]()