INTRODUCTION
Cycling of sulfur compounds is a ubiquitous process in marine sediments that supports a range of microbial metabolic strategies. The occurrence of sulfur over a wide range of oxidation states (–2 to +6) allows sulfur species to serve as both electron acceptors and electron donors. In reduced form as sulfide (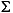 H2S = H2S(aq) + HS–), sulfur is also an important sink for reactive iron (Berner, 1971; Goldhaber and Kaplan, 1974). The reduction of sulfate to sulfide is by far the most important pathway for sedimentary organic matter oxidation in anoxic marine sediments, and there is increasing evidence that anaerobic oxidation of methane controls microbial sulfate reduction (MSR) in many marine systems (Niewöhner et al., 1998; Aharon and Fu, 2000; Hensen et al., 2003; D'Hondt, Jørgensen, Miller, et al., 2003).
H2S = H2S(aq) + HS–), sulfur is also an important sink for reactive iron (Berner, 1971; Goldhaber and Kaplan, 1974). The reduction of sulfate to sulfide is by far the most important pathway for sedimentary organic matter oxidation in anoxic marine sediments, and there is increasing evidence that anaerobic oxidation of methane controls microbial sulfate reduction (MSR) in many marine systems (Niewöhner et al., 1998; Aharon and Fu, 2000; Hensen et al., 2003; D'Hondt, Jørgensen, Miller, et al., 2003).
The stable isotope ratios of oxygen (18O/16O) and sulfur (34S/32S) in dissolved sulfate may provide information about specific biogeochemical pathways for sulfur in marine sediments, such as bacterial sulfate reduction, sulfide oxidation, sulfur disproportionation, and transport processes of sulfur species (Rees, 1973; Goldhaber and Kaplan, 1980; Zak et al., 1980; Canfield et al., 1998; Böttcher et al., 1998a, 1998b, 1999, 2001, 2004; Jørgensen et al., 2004; Brunner et al., 2005). Sulfate containing the lighter isotope 32S is utilized preferentially during MSR, resulting in an enrichment of 34S in the residual pore water sulfate. In seawater-derived pore waters, an enrichment of 18O is also observed during ongoing MSR (Zak et al., 1980; Böttcher et al., 1998a, 1998b, 1999, 2004). The magnitudes of isotope fractionation accompanying MSR and the degree of heavy isotope enrichment in residual sulfate seem to depend on a number of environmental factors, including temperature, sulfate concentration, microbial community composition, the rate of sulfate reduction, and the type and amount of sedimentary organic matter being oxidized (Jørgensen, 1979; Berner, 1980; Boudreau and Westrich, 1984; Aharon and Fu, 2000). Isotopic fractionation of oxygen and sulfur in residual sulfate during MSR has been measured in pure laboratory cultures, sediment incubations, and natural systems and ranges from 4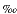 to 29 for oxygen and from 4 to 46 for sulfur (e.g., Kaplan et al., 1963; Lloyd, 1967, 1968; Kemp and Thode, 1968; Mizutani and Rafter, 1973; Chambers and Trudinger, 1979; Fritz et al., 1989; Ku et al., 1999; Böttcher et al. 2001; Aharon and Fu, 2000; Mandernack et al., 2003). Whereas sulfur isotope fractionation upon dissimilatory sulfate reduction is usually interpreted as primarily kinetic in nature (e.g., Kaplan and Rittenberg, 1964; Rees, 1973), an intracellular isotope exchange between water and sulfur intermediates is proposed to lead to a partial oxygen isotope exchange between overall extracellular sulfate and pore waters (Fritz et al., 1989; Böttcher et al., 1998a, 1999; Aharon and Fu, 2000). Under oxygen isotope exchange equilibrium conditions, dissolved sulfate should be enriched in 18O compared to Vienna-standard mean ocean water (V-SMOW) by more than 30 (Böttcher et al., 1998a).
to 29 for oxygen and from 4 to 46 for sulfur (e.g., Kaplan et al., 1963; Lloyd, 1967, 1968; Kemp and Thode, 1968; Mizutani and Rafter, 1973; Chambers and Trudinger, 1979; Fritz et al., 1989; Ku et al., 1999; Böttcher et al. 2001; Aharon and Fu, 2000; Mandernack et al., 2003). Whereas sulfur isotope fractionation upon dissimilatory sulfate reduction is usually interpreted as primarily kinetic in nature (e.g., Kaplan and Rittenberg, 1964; Rees, 1973), an intracellular isotope exchange between water and sulfur intermediates is proposed to lead to a partial oxygen isotope exchange between overall extracellular sulfate and pore waters (Fritz et al., 1989; Böttcher et al., 1998a, 1999; Aharon and Fu, 2000). Under oxygen isotope exchange equilibrium conditions, dissolved sulfate should be enriched in 18O compared to Vienna-standard mean ocean water (V-SMOW) by more than 30 (Böttcher et al., 1998a).
Much of the sulfide produced during dissimilatory MSR in marine sediments is oxidized back to sulfate by a variety of biological and abiotic pathways (Jørgensen, 1990; Goldhaber and Kaplan, 1980; van Stempvoort and Krouse, 1994; Canfield and Thamdrup, 1994; Böttcher and Thamdrup, 2001), and sulfate produced by oxidation of sulfide may have variable  18O values reflecting the nature and complexity of the abiotic and biological oxidation pathways and relative contributions from different oxidants (water and dissolved oxygen) (Taylor et al., 1984; Böttcher et al., 2001; Brunner et al., 2005). These pathways often include the production of intermediate sulfur species such as elemental sulfur and thiosulfate, which can undergo further bacterial disproportionation reactions that may lead to further fractionations of both sulfur and oxygen isotopes in secondary sulfate (Canfield and Thamdrup, 1994; Canfield et al., 1998; Cypionka et al., 1998; Habicht et al., 1998; Böttcher et al., 2001, 2004; Böttcher and Thamdrup, 2001). Under circumneutral and low-temperature sedimentary conditions, an abiotic oxygen isotope exchange between water and sulfate does not take place (Lloyd, 1967, 1968; Zak et al., 1980). Therefore, changes in 18O values of sulfate under anoxic conditions provide clear evidence for the enzyme-catalyzed processes. Thus, the stable isotope ratio of oxygen in dissolved sulfate may be a valuable indicator for tracing microbial activity and sulfur transformations by a deep biosphere in marine sediments.
18O values reflecting the nature and complexity of the abiotic and biological oxidation pathways and relative contributions from different oxidants (water and dissolved oxygen) (Taylor et al., 1984; Böttcher et al., 2001; Brunner et al., 2005). These pathways often include the production of intermediate sulfur species such as elemental sulfur and thiosulfate, which can undergo further bacterial disproportionation reactions that may lead to further fractionations of both sulfur and oxygen isotopes in secondary sulfate (Canfield and Thamdrup, 1994; Canfield et al., 1998; Cypionka et al., 1998; Habicht et al., 1998; Böttcher et al., 2001, 2004; Böttcher and Thamdrup, 2001). Under circumneutral and low-temperature sedimentary conditions, an abiotic oxygen isotope exchange between water and sulfate does not take place (Lloyd, 1967, 1968; Zak et al., 1980). Therefore, changes in 18O values of sulfate under anoxic conditions provide clear evidence for the enzyme-catalyzed processes. Thus, the stable isotope ratio of oxygen in dissolved sulfate may be a valuable indicator for tracing microbial activity and sulfur transformations by a deep biosphere in marine sediments.
Only a few studies on marine pore water systems have included the analysis of the stable oxygen isotopic composition of dissolved sulfate to date (Zak et al., 1980; Böttcher et al., 1998a, 1998b, 1999; Bottrell et al., 2000), and most have focused on shallow pore water systems (Pierre, 1985; Böttcher et al., 1998b; Ku et al., 1999; Aharon and Fu, 2000). The sites drilled during Ocean Drilling Program (ODP) Leg 201 provide a unique opportunity to study oxygen isotope fractionation by the deep biosphere in pore waters of deep sediment sections averaging several hundred and as much as ~400 meters below sea floor (mbsf) and cover a wide range of water depths and sedimentary conditions (e.g., organic carbon content and sedimentation rate).
