 Cryptocarpium
ornatum (Ehrenberg)Cryptocarpium
ornatum (Ehrenberg)
Cryptocarpium
ornatum (Ehrenberg)Cryptocarpium
ornatum (Ehrenberg)Cryptoprora ornata Ehrenberg, 1873, p.222; 1875, pl.5, fig.8
Cryptocarpium ornatum (Ehrenberg) Sanfilippo and Riedel, 1992, p.6, pl.2, figs.18-20
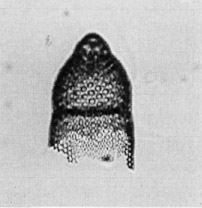
Three-segmented pterocorythid with a cephalis that, in early forms, is as prominent as in a typical pterocorythid to as indistinct as in a typical carpocaniid in late forms. Thorax hemispherical, sometimes with a thickened wall apically. In these specimens the cephalis appears flattened, and the collar stricture is indistinct. Abdomen usually cylindrical or slightly expanded distally. The abdominal lattice is similar to that of the thorax, with pores closely spaced in longitudinal rows, not separated by ridges. The termination is undifferentiated. Longitudinal ridges between abdominal pore rows have been observed in well developed late forms (Sanfilippo, unpubl. data).
Total length 90-175 mm; of cephalothorax 70-105 mm; maximum width of thorax 65-100 mm (Sanfilippo et al., 1985).
C. ornatum is distinguished from Cryptocarpium azyx as indicated under that species. It is distinguished from Carpocanopsis cingulata, from which it is far separated stratigraphically, by the thinner thoracic wall and subcylindrical or distally expanding abdomen. Near the top of its range, Cryptocarpium ornatum is accompanied by an apparently related form which differs in having longitudinal ridges between rows of thoracic pores, and the thorax tending to become narrower distally. C. ornatum grades from a narrow, non-tuberose precursor of Theocyrtis tuberosa, the line of demarcation being drawn at the point at which the apical horn is lost (Sanfilippo et al., 1985).
See also Sanfilippo and Riedel, 1992, pl.2, fig. 17 (non-tuberose precursor of T. tuberosa) and pl.2, fig.21 (Cryptocarpium azyx).
This three-segmented form has abdominal lattice similar to that of the thorax, with pores closely spaced in longitudinal rows, not separated by ridges. The most variable characters are the overall lengths and the degree of prominence of the cephalis. In early forms the cephalis is as prominent as in a typical pterocorythid, and in late forms it is as distinct as in a typical carpocaniid. In some specimens the abdomen slightly expands distally, and there is never a differentiated termination (Sanfilippo et al., 1985).
C. ornatum is found in late middle Eocene to earliest Oligocene assemblages from tropical localities in all three oceans, and as far south as DSDP Site 283, near Tasmania. Its morphotypic first appearance is within the Podocyrtis mitra Zone and its morphotypic last appearance is approximately synchronous with the lower limit of the Theocyrtis tuberosa Zone.
This species originated from Cryptocarpium sp., an ancestor in common with that of the genus Albatrossidium. The distinct three-lobed cephalic structure originates in the genus Cryptocarpium in the middle Paleocene from simple three-segmented theoperids with an apical bar that gives off a pair of mitral arches near its junction with the shell wall. In the transition from Cryptocarpium sp. not only was the horn lost, but the cephalis eventually became flatter and the collar stricture indistinct. The late forms thus have a reduced cephalis similar to that of the carpocaniids. However, the carpocaniids of the Theocyrtis tuberosa Zone are not forms with pores pronouncedly aligned longitudinally and possessing post-thoracic structure (as in Carpocanopsis cingulata and Cryptocarpium ornatum), but instead are simple Carpocanistrum spp. with pores not pronouncedly aligned except near the top of the zone, and having the thorax terminated by a plain peristome (as illustrated by Riedel and Sanfilippo, 1971, pl.3D, figs.6-8). Cryptocarpium ornatum gave rise to C. azyx in the late Eocene and became extinct at the bottom of the Theocyrtis tuberosa Zone.
Additional illustrations can be found in Sanfilippo and Riedel, 1973, pl.35, figs.3-4.
For generic level taxonomy see Sanfilippo and Riedel, 1992.
![]()