 Dictyoprora
mongolfieri (Ehrenberg)Dictyoprora
mongolfieri (Ehrenberg)
Dictyoprora
mongolfieri (Ehrenberg)Dictyoprora
mongolfieri (Ehrenberg)Eucyrtidium mongolfieri Ehrenberg, 1854, pl.36, fig.18, B lower; 1873, p.230
Dictyoprora mongolfieri (Ehrenberg), Nigrini, 1977, p.250, pl.4, fig.7
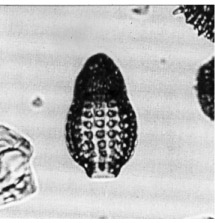
Cephalis hemispherical, with circular pores, and with an inconspicuous lateral tubule lying against the upper part of the thoracic wall. Collar stricture indistinct. Thorax slightly inflated annular, with circular pores inclined obliquely upwards and inward as they traverse the thick wall. Lumbar stricture distinct. Abdomen inflated barrel-shaped, constricted distally to a short cylindrical, hyaline peristome. Abdominal pores strictly in longitudinal and transverse rows, the longitudinal rows separated by ridges (Riedel and Sanfilippo, 1970).
Total length of shell 125-160 µm; maximum breadth 75-100 µm (Nigrini, 1977).
D. mongolfieri is distinguished from all other species of Dictyoprora by the abdominal pores being in strict longitudinal and transverse rows. It differs from D. armadillo (Ehrenberg, 1873, p.225) in lacking an apical horn and having an ellipsoidal rather than pyriform abdomen.This species has been widely used in stratigraphy because it is relatively easily recognized, occurs fairly abundantly throughout its range, and can be recognized in assemblages that are not very well preserved. Its [distinction] from other artostrobiid species [of the D. amphora (Haeckel) group Foreman, 1973] is not completely satisfactory in some assemblages, where specimens with abdominal pores strictly aligned longitudinally [co-occur with transitional forms] having abdominal pores quincuncially arranged. Uncertainties are best avoided by not admitting specimens departing to any degree from strictly longitudinal and transverse alignment of the pores. Even so, there is a problem in recognizing the [lowermost] limit of occurrence of D. mongolfieri. Because it is a morphotypic rather than an evolutionary limit, single specimens justify its being recorded as present, and their detection in [the earliest] assemblages depends on the amount of effort spent in searching. This difficulty cannot be avoided until the evolutionary origin of the species is satisfactorily determined (Sanfilippo et al., 1985)
Throughout its stratigraphic range, D. mongolfieri varies only slightly in the lumbar stricture being more or less distinct, depending on the degree of inflation of the abdomen. The abdominal pores are always strictly aligned rectangularly in longitudinal and transverse rows, with the longitudinal rows separated by ridges. Late forms tend to be a little wider than earlier forms (75-100 µm as compared to 65-90 µm), with somewhat more pores on a half-diameter of the abdomen (9-11 as compared to 7-9). The simple, short, hyaline peristome also undergoes a change toward the end of the range, to an undulating margin or three smooth shovel-shaped feet in some specimens (Sanfilippo et al., 1985).
D. mongolfieri is common at all tropical (30°S-30°N) localities from which sediments of middle middle Eocene to early early Oligocene age are available. The morphotypic first appearance of this species defines the base of the Dictyoprora mongolfieri Zone. Its morphotypic last appearance is approximately synchronous with the lower limit of the Theocyrtis tuberosa Zone.
D. mongolfieri might have arisen from the D. amphora group, but this group is very varied and has a similar stratigraphic range to D. mongolfieri. It left no known descendants.
Additional illustrations can be found in Riedel and Sanfilippo, 1971, pl.3E, fig.13; Nigrini, 1974, pl.lM, figs.7-10, pl.2E, fig.9.
![]()