DISCUSSION
A good paleoclimatic record has been obtained from the planktonic foraminiferal assemblage of both Sites 975 and 976 in the Western Mediterranean. The close correspondence between paleoclimatic trends and the oxygen-isotope record suggests a strong control of the temperature in faunal planktonic distributions. Also, rock magnetic parameters show a correlation with the paleoclimatic record, indicating that climate exerts a major role on the magnetic properties of these sediments.
By using the ages of well-identified points (bioevents, oxygen-isotope stage boundaries, and climatic events, Blake Event), it has been possible to reconstruct a detailed sedimentation rate for these sites (Fig. 11). A significant difference exists between the Balearic Basin and the Alboran Sea, which reflects the different amount in the supply of terrigenous material between the two sites. In the Balearic Basin we have a quite constant sedimentation rate of about 7 cm/k.y. from the Holocene back to oxygen-isotope stage 4. Corresponding to the beginning of oxygen-isotope stage 5, there is an apparent decrease in the sedimentation rate that is a result of a gap recognized between the end of the oxygen-isotope stage 4 and substage 5c (Fig. 5). In particular, the presence of a "sapropel-like" planktonic foraminiferal assemblage corresponding to the end of this hiatus suggests that the missing record spans the time interval beginning at the end of the oxygen-isotope stage 4 (about 73.9 k.y.; Martinson et al., 1987) and the sapropel S4 (102 k.y.; Lourens et al., 1996).
It is noteworthy that a significant increase in the sedimentation rate occurs during the Eemian (substage 5e). At Site 975 the sediments also exhibit a low organic carbon content (Fig. 4) that above 9.5 m of depth is almost constantly below the values recorded in normal open-marine environments (0.3%; Stein 1991).
At Site 976, the sedimentation rate ranges between 34 and 50 cm/k.y. Higher sedimentation rates occur for the last two oxygen-isotope stages. The total organic carbon measured at this site exceeded the values of 0.5% in most of the samples (Fig. 4), which is distinctly higher than those observed in open-marine environments. The carbon/nitrogen ratio (C/N) is quite constant with values ranging between 4 and 11, indicating the predominance of marine organic matter in the Alboran Sea (Fig. 4). Furthermore, the organic-carbon/sulfur ratio indicates that several intervals were deposited during anoxic conditions.
The TOC profile exhibits a trend that appears correlated with the climatic variations. High/low organic carbon content corresponds to warm/cold periods (Fig. 4). Nevertheless a couple of intervals show a different feature. In fact, the Termination I and II are both characterized by peaks in the TOC values, whereas the Eemian substage yields quite low values. The observed oscillations cannot be related to changes in the sedimentation rate, which would enhance the preservation of the organic matter (Stein, 1991). Therefore, the variations in the organic matter content correspond to changes in the primary productivity. A parallelism between the TOC values and the N. dutertrei distribution supports this observation (Fig. 12). The high abundance of this species reflects the presence of a Deep Chlorophyll Maximum (DCM; Thunell, 1979; Fairbanks et al., 1982; Rohling and Gieskes, 1989; Pujol and Vergnaud-Grazzini, 1995).
It is noteworthy that both Termination I (including the Younger Dryas) and II are characterized by peak values in the C/N ratio with values that suggest a possible increasing contribution of terrestrial organic matter during these periods (Fig. 4).
Possible Sources of the Magnetic Minerals
The fluctuations in magnetic concentration and grain size reflect changes in the types of magnetic minerals supplied to the sediments, as well as the nature of depositional paleoenvironments. In general, the magnetic concentration is low in biogenic sediments and high in terrigenous materials. Exceptions to this feature often occur in marine sediments because (1) dilution by other sedimentary components; (2) increasing distance from sources of magnetic minerals; (3) diagenetic processes; and (4) authigenic formation of bacterial magnetite.
The magnetic properties measured in the samples from Site 974, 975, and 976 exhibit differences that are related to either different sources of the magnetic minerals or to different environmental conditions of the basins. At all the sites the dominant magnetic mineral is magnetite. S-ratio values suggest that a relative importance of antiferromagnetic minerals occurs only during the glacial stages at Site 975 and in the intervals of reductive diagenesis observed at Site 976. In the former site, the antiferromagnetic minerals present within the sediments are probably related to an increasing contribution of the aeolian fraction (Saharan dust) during these cold stages. The highest concentration of magnetic minerals occurs in the Tyrrhenian Sea and the possible source of the relatively coarse-grained magnetite in this area can be identified with the volcanic provinces of central and southern Italy (McCoy and Cornell, 1990). In the Balearic Basin (Site 975), the KARM profile exhibits a correlation with glacial and interglacial stages (Fig. 7), which would suggest that bacterial magnetite is the most likely source for these sediments. In the Alboran Sea, the ferrimagnetic minerals within the sediments are likely supplied by terrigenous material. Site 976 is located in shallow water with respect to the other sites, where very probably the source area for the magnetic materials lies in the southern Spanish coast. These observations are supported by the plot of ARM/SIRM vs. IRM100/SIRM shown in Figure 13. This plot reflects the contribution of SSD (high ARM/SIRM values) with respect to the ferrimagnetic:antiferromagnetic ratio (increasing values of IRM100/SIRM). The samples from Site 975 exhibit the highest ARM/SIRM value within a range that includes the values obtained for deep-sea sediments considered to have magnetic properties dominated by bacterial magnetosomes (Oldfield, 1994). An unequivocal interpretation of these data would require microscope observations of the typical structure of bacterial magnetite and will be the subject of a future investigation.
At Site 976 concentration (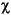 , SIRM, ARM), grain-size (Karm/, SIRM/, ARM/SIRM) and coercivity (S-ratio, B0cr) parameters show a similar trend, implying that they are controlled by a unique factor. This factor is a process of reductive diagenesis related to bacterial degradation of organic matter. The organic-carbon/sulfur relationship clearly shows the difference between this site and the site drilled in the Balearic Basin (Fig. 10).
, SIRM, ARM), grain-size (Karm/, SIRM/, ARM/SIRM) and coercivity (S-ratio, B0cr) parameters show a similar trend, implying that they are controlled by a unique factor. This factor is a process of reductive diagenesis related to bacterial degradation of organic matter. The organic-carbon/sulfur relationship clearly shows the difference between this site and the site drilled in the Balearic Basin (Fig. 10).
The quantitative planktonic distribution allows one to reconstruct a detailed paleoclimatic trend for the western Mediterranean during the late Quaternary. The microfaunal assemblage at both Sites 975 and 976 document the presence of the Holocene in the uppermost interval. At Site 976, which is characterized by a quite high sedimentation rate, the abundance at the top of the core of G. bulloides and G. inflata suggests that it represents the present-day assemblages. In fact, at present, the Alboran Sea is characterized by the dominance of G. inflata during winter time (Deveaux, 1985) and by G. bulloides during summer season (Cifelli, 1974).
The reconstructed climatic trend is characterized by cold-temperate conditions with high values during the Holocene time and Eemian interval (oxygen-isotope substage 5e). The two oxygen-isotope boundaries 6/5 and 2/1 corresponding respectively to Termination II and I, are characterized by the same pattern: the passage between two extreme climatic conditions (cold and warm) is rapid and very well documented. At Site 975, a higher sedimentation rate during the Eemian substage (45cm/k.y.) documents a detailed paleoclimatic record. Significant fluctuations appear recorded during this time interval suggesting that it is divided into three substages, as also observed in North Atlantic cores by Fronval and Jansen (1996).
The paleoclimatic record shows a cyclicity that is also well recorded by magnetic parameters. This trend is similar to the rapid oxygen-isotope oscillations observed in the GRIP ice-core (Dansgaard et al., 1984). Several abrupt colder episodes are characterized by the dominance of cold species with a peak in frequency of N. pachyderma (left-coiling) and/or G. quinqueloba. This association, together with the minimum in the number of planktonic foraminifers is exactly the same biological record observed in the Heinrich events documented in the North Atlantic sediments (Bond et al., 1993; Maslin et al., 1995). Some of these events can also be identified in the magnetic parameter profiles. All this evidence suggests that the oceanographic setting of Site 976 is strongly influenced by the water-mass exchange across the Gibraltar sill.
Occurrence of Sapropels in the Western Mediterranean
Sapropel layers were identified at both sites. At Site 975 the considered time interval includes sapropels S1-S9 in the Eastern Mediterranean Basin, and some of them have been identified within the cores. Most of these layers (S5, S6, S7, S9; Fig. 4) correspond to well-identified dark intervals with rock magnetic properties typical of anoxic environments. As discussed in the "Results" section, this chapter, the sapropels S3 and S4 belong to an interval that is missing in the sequence (Fig. 5).
At Site 976 the sapropel identification is not straightforward. Neither color nor organic-carbon content can be used as discriminant factors. No main changes occur in the sediment color and the organic-carbon profile exhibits several peaks scattered along the core. The site was anoxic during most of the time interval studied, and this makes it difficult to recognize the sapropels by using "anoxic features." The planktonic microfossil record has been the first and discriminant criteria for sapropel identification.
Several studies have shown that an unusual planktonic foraminiferal fauna characterizes the sapropel with peaks in frequencies of G. ruber, G. bulloides, O. universa, and, in many levels, of N. dutertrei (Cita et al., 1977; Vergnaud-Grazzini et al., 1977; Thunell and Lohmann, 1979; Muerdter and Kenneth, 1983/84). In most of these layers, the planktonic assemblage (abundance of N. dutertrei) and the oxygen-isotope depletion suggest that the surface-water salinity was relatively low (Cita et al., 1977; Vergnaud-Grazzini et al., 1977; Williams and Thunell, 1979; Muerdter, 1984; Tang and Stott, 1993). We have noted that it is possible to discriminate the sapropels on the basis of the quantitative and qualitative variations in planktonic foraminifer assemblage, either in warm or cold intervals. The sapropel layers deposited during warm intervals are characterized by peaks in frequencies of G. ruber and occurrence of G. ruber var. rosea, while abundance of N. dutertrei characterizes the sapropel layers deposited in cold intervals. This feature is also typical of the sediments from the western Mediterranean as shown in Figure 12. Both these species inhabit surface waters and are well documented in the literature to be related to low salinity (Bé and Tolderlund, 1971; Cita et al., 1977; Fairbanks et al., 1982; Rohling and Gieskes, 1989; Tang and Stott, 1993). Qualitative evidence shows the presence of several species with very thin tests and large pores, which can be easily detected even by optical microscope (see
fig. 9 in Borsetti et al., 1995). Together with the presence of organic matter, which adheres to the test (as reported in
fig. 7 of Cita et al., 1996) or pyritization processes, there are characteristic features of planktonic assemblages observed within sapropel layers found in the eastern Mediterranean Sea.
The high microfaunal commonality with the assemblage documented during the sapropel layer deposition in the Eastern Mediterranean Sea, combined with increasing organic-carbon content, magnetic measurements, and the stratigraphic position, support our interpretation.
The stratigraphic position of the recognized sapropel layers at Sites 975 and 976, together with the associated total organic carbon content, is listed in Table 5 and Table 6. The estimated age for the recognized sapropel layers at both the sites 975 and 976 (reported in Table 5 and Table 6) is in agreement with the age obtained by astronomical calibration for the same sapropels in the eastern Mediterranean (Lourens et al., 1996).
Finding sapropel layers in the western Mediterranean agrees with the oceanographic setting of the basin as shown by the presence of the Levantine Intermediate Water in the Gibraltar Sill (Loubere, 1987).
