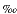 ) relative to the Peedee belemnite (PDB) standard.
) relative to the Peedee belemnite (PDB) standard.
The late Quaternary sediments at Site 1002 are generally dominated by terrigenous sediment components with variable biogenic contributions of nannofossils, diatoms, foraminifers (both planktonic and benthic), and pteropods. Throughout much of the sequence, the presence of aragonite pteropods is indicative of the generally excellent preservation of calcareous microfossils. Much of the sediment at Site 1002 is laminated, indicating deposition under nearly anoxic conditions (Fig. 2). Nevertheless, significant subsurface intervals showing clear evidence of bioturbation testify to a history of oscillation between oxic and anoxic environments in the deep basin. Because no major changes in bulk sediment composition occur over the interval drilled, the sediments at Site 1002 were assigned to one formal lithologic unit and the eight subunits illustrated in Figure 2 and Figure 3.
The sedimentary sequence at Site 1002 essentially duplicates the section recovered at DSDP Site 147 at virtually the same location in the central Cariaco Basin. However, in terms of recovery and core quality, there is almost no comparison between the two drilling efforts. Whereas Site 147 cores were so badly disturbed that subsurface laminae were only rarely found preserved, the APC and XCB cores from Site 1002 reveal a sequence that is intermittently laminated over much of its length.
Although cores from DSDP Site 147 were disturbed, thin layers of distinct yellowish brown and bluish gray clays similar to what were found near the surface were also identified at several deeper levels in the section (Fig. 3). The DSDP Leg 15 scientists (Edgars, Saunders, et al., 1973) postulated that these deeper clays represented earlier periods of oxic conditions comparable to those recorded during the LGM. They further suggested that sedimentation patterns in the Cariaco Basin followed a "rhythmic" alternation (Fig. 3) between periods of oxic and anoxic conditions that were related to the glacial-interglacial cycles of the late Quaternary. The lack of good biostratigraphic datums over the time interval represented by the sediments, however, effectively prevented their association of these "rhythms" with specific climatic events.
As already noted, the tentative identification of Emiliania huxleyi in core catchers at the base of Hole 1002C, a species that, under shipboard conditions, is difficult to distinguish from small specimens of Gephyrocapsa with etched central area bridges, initially led us to believe that the sedimentary sequence recovered during Leg 165 was no older than 248 ka. Indeed, the great visual similarity between the very colorful and distinctive bioturbated clays of Subunits IB and IG, and the abrupt transition to the laminated, diatom-rich anoxic sediments of overlying Subunits IA and IF, respectively, led to speculation (Shipboard Scientific Party, 1997b) that the Subunit IG-IF transition at ~98 mbsf in Site 1002 recorded the rapid deglaciation and sea level rise at ~128 ka associated with the change from marine isotope Stage (MIS) 6 to MIS 5. Shipboard observations of Globorotalia tumida flexuosa in the core catcher of Core 165-1002C-8H, but no higher, tended to support this hypothesis because the last occurrence datum for this taxon (80 ka) approximates the end of MIS 5. Our working model that the Subunit IG/IF boundary at ~98 mbsf corresponded to the MIS 6/5 deglaciation appeared to be further consistent with estimates based on sedimentation rates derived from an assumed sequence age of <248 ka, and with knowledge that the lithologically identical Subunit IB/IA transition was already known (e.g., Peterson et al., 1991) to coincide with the most recent deglaciation (MIS 2/1). It also seemed to fit the Leg 15 proposition that the repetitive sequence of lithologies in DSDP Site 147 reflected sedimentation cycles somehow linked to climate (Fig. 3). Postcruise efforts, however, have shown this preliminary age model to be in error as new data from shore-based stable isotope stratigraphy and biostratigraphic studies have become available.
Samples for stable isotope analysis were initially taken at a frequency of one sample per 1.5-m section from all cores in Hole 1002C except Core 165-1002C-9H. Much of Core 9H was disturbed during recovery when the wireline broke and the APC barrel fell back to the bottom of the hole. To avoid including samples from this disturbed interval (~76 to 84.4 mbsf), samples were taken from Hole 1002D and spliced into the sequence from Hole 1002C. Once a preliminary stratigraphy was available at this resolution, additional samples with an average spacing of 10-30 cm and 40-60 cm were obtained from Hole 1002C that spanned the intervals that were determined to include MISs 2-4 and MIS 5, respectively. Data presented here were obtained from measurements on specimens of the shallow, mixed-layer dwelling planktonic foraminifer, Globigerinoides ruber (250-355 µm). Only specimens of the white morphotype of G. ruber were analyzed because Lin et al. (1997) found systematic isotopic differences between the white and pink forms of this taxon in the Cariaco Basin. All isotope analyses were made on a Finnigan-MAT 251 mass spectrometer in the Stable Isotope Laboratory of the Rosenstiel School at the University of Miami. Isotope data are reported
(Table 1) in units of parts per thousand () relative to the Peedee belemnite (PDB) standard.
The  18O data for G. ruber in Hole 1002C are plotted in Figure 4 together with selected biostratigraphic datums. Because of gas expansion in the core liners that occurred as cores were brought to the surface, recovery in the APC-cored Hole 1002C averaged 109.1%. To correct for this, gas voids in excess of 2 cm thick were subtracted from the sediment column and a linear compression was applied to each 1.5-m core section so that the compressed core length was equal to the actual interval cored (Piper and Flood, 1997). Hence, data in
Figure 4 are plotted vs. this "corrected mbsf."
18O data for G. ruber in Hole 1002C are plotted in Figure 4 together with selected biostratigraphic datums. Because of gas expansion in the core liners that occurred as cores were brought to the surface, recovery in the APC-cored Hole 1002C averaged 109.1%. To correct for this, gas voids in excess of 2 cm thick were subtracted from the sediment column and a linear compression was applied to each 1.5-m core section so that the compressed core length was equal to the actual interval cored (Piper and Flood, 1997). Hence, data in
Figure 4 are plotted vs. this "corrected mbsf."
The 18O record for Hole 1002C can be easily matched to the well-known late Quaternary signal (e.g., Emiliani, 1955) down to about the MIS 14/13 boundary. The identification of isotope stages is consistent with a refined biostratigraphic framework largely anchored by the classic Ericson zonation scheme (Ericson et al., 1961; Ericson and Wollin, 1968). The latter is based on recognition of ecostratigraphic zones in the tropical Atlantic Ocean defined by the presence or absence of Globorotalia menardii and Globorotalia tumida. Also identified in Hole 1002C are the revised FO of the calcareous nannofossil Emiliania huxleyi, the LO of Pseudoemiliania lacunosa in MIS 12, and the disappearance of the planktonic foraminifer Pulleniatina obliquiloculata within the Ericson Y zone (YP. obliq.). The regional disappearance of P. obliquiloculata is time-transgressive within the western tropical Atlantic and Caribbean and has been placed between 44 and 36 ka by Prell and Damuth (1978). Biostratigraphic datums used to constrain the 18O stratigraphy are listed in
Table 2.
Figure 5 shows an expanded view of the more detailed record presently available for MIS 1-6. The overall glacial-interglacial amplitude of the G. ruber 18O signal, both here and in the remainder of the record
(Fig. 4) is large, typically well in excess of 2. This amplitude is similar to that previously reported by Lin et al. (1997) for this taxon for the last deglaciation. In theory, residual differences between the amplitude of the Cariaco Basin 18O signal and the change in mean ocean 18O composition expected from meltwater addition during deglaciation (~1.2-1.3; Fairbanks, 1989) should be attributable to local temperature and/or salinity effects. If solely ascribed to temperature, the residuals could represent as much as 4°-6°C of cooling of glacial surface waters over the Cariaco Basin, assuming a temperature:18O relationship of 0.22 per 1°C (Epstein et al., 1953). In an earlier analysis of planktonic foraminiferal assemblages, Peterson et al. (1991) found no clear evidence of glacial cooling in the Cariaco Basin. However, these authors also noted the selective exclusion of deeper dwelling taxa from the basin during sea-level lowstands, making quantitative SST estimates (i.e., by transfer functions) highly suspect. More recently, Lin et al. (1997) interpreted 18O data from G. ruber for the last glacial and reduced 18O differences among the multiple taxa analyzed to indicate glacial cooling over the Cariaco Basin of up to ~4°C. Herbert and Schuffert (Chap. 16, this volume), on the other hand, provide intriguing alkenone paleotemperature data from Hole 1002C sediments which show little cooling, if any, during the LGM and during MIS 6, but variations of up to 4°C in the earlier part of MIS 3, in MIS 4, and the later part of MIS 5. Interpreted at face value, these data would suggest that salinity variations must, at times, have played a substantial role in producing the large amplitude of the Cariaco Basin isotopic signal.
As will be discussed later in more detail, the Cariaco Basin would have been considerably more isolated during maximum sea level lowstands, likely resulting in higher salinities through basin restriction and increased evaporation. For the LGM, where alkenone data suggest little temperature change, Herbert and Schuffert (this volume) conclude that a salinity increase of ~2 would be required to explain the ~1 residual enrichment in foraminiferal 18O. Support for such a salinity increase can indeed be found in foraminiferal data. Peterson et al. (1991) noted that higher glacial salinities in the Cariaco Basin could at least partially explain the dominance of G. ruber in LGM sediments. In addition, preliminary benthic foraminiferal census data from Hole 1002C in at least some intervals of glacial Stages 2-4 show that populations are dominated by members of the miliolid genus Pyrgo, which are high-Mg calcite, and by the aragonitic genus Hoeglundina. Both taxa are common in high salinity, carbonate-saturated environments today such as the Red Sea and Persian Gulf (e.g., Halicz and Reiss, 1981; Locke and Thunell, 1988).
Superimposed on the orbital-scale variability of the MIS 1 to 6 interval is clear evidence for millennial and submillennial variability in 18O (Fig.
5) that can probably also be attributed to local temperature and/or salinity effects. Similar high-frequency oscillations in a planktonic foraminiferal 18O record (EW9209 JPC) from the open western equatorial Atlantic have been observed by Curry and Oppo (1997). The amplitude of the high-frequency 18O oscillations at that location, interpreted by Curry and Oppo (1997) as a temperature signal, is ~0.6 and are superimposed on a glacial-interglacial amplitude of ~2.1. Measured amplitudes of the high-frequency oscillations at Site 1002 are larger, but may be a function of the resolution afforded by the much higher deposition rates at this site. Although it is tempting to speculate that low isotopic values most evident in sediments of MIS 3 in the Cariaco Basin
(Fig. 5) can be correlated to individual high-frequency events in the Curry and Oppo (1997) record from EW9209 JPC, and, in turn, to the well-known warm interstadial events in the Greenland ice core records (e.g., GRIP Members, 1993), the age control at Site 1002 is not yet robust enough to attempt this match. Furthermore, salinity effects related to potential freshwater discharge into the Cariaco Basin must also be carefully evaluated. Clear evidence of a major freshwater event in the Cariaco Basin at ~14 ka has already been documented by Lin et al. (1997), suggesting the potential for similar events earlier in the record. The question of abrupt events and high-frequency variability in 18O and lithologic records of Site 1002 will be more fully evaluated elsewhere.
Comparison of the long 18O record in Hole 1002C with the downcore distribution of lithologic subunits leads to other interesting observations
(Fig. 4). Sediments of Subunit IG, consisting of colorful interbedded light bluish gray and pale brown clays, are visually and lithologically most similar to the clays of Subunit IB that were deposited in the latter stages of MIS 2 (LGM). As noted above, it was this visual similarity that largely contributed to earlier speculation (Shipboard Scientific Party, 1997b) that the Subunit IG/IF boundary corresponded to the sharp deglaciation (Termination II) associated with the MIS 6/5 transition. The 18O stratigraphy for Hole 1002C, however, clearly indicates that Subunit IG was deposited much earlier, near the end of MIS 10, and that the MIS 6-5 transition actually falls at an expansion-corrected depth of ~46 mbsf. Why clays associated with the termination of glacial MIS 10 are the only other sediments at Site 1002 visually identical to the distinct yellow and gray clays of the LGM is not clear. Whatever the reason, the simple model of sedimentary "rhythms" first proposed for DSDP Site 147
(Fig. 3) is clearly more complex than originally envisioned, with individual rhythms encompassing more than one complete glacial-interglacial cycle.
A first-order correlation of the Hole 1002 18O record to the SPECMAP composite standard for the late Quaternary (Imbrie et al., 1984) is shown in
Figure 6. Although resolution of the Hole 1002C record is relatively low below MIS 5 because of the wide sampling interval (1 sample/section) currently used, filtered comparisons of the Site 1002 and SPECMAP 18O records in the orbital bands of precession and obliquity indicate that the two records are well matched in the time domain
(Fig. 7). The present age model (Fig.
8) indicates a basal age of ~580 ka for the recovered Cariaco Basin sequence, yielding an average sedimentation rate of ~35 cm/k.y. (350 m/m.y.). This rate is similar to previous estimates of sedimentation for the uppermost sequence (Peterson et al., 1991; Hughen et al., 1996b; Lin et al., 1997).
Diatom-rich sediments are characteristic of the well-laminated lower portion of Subunit IA, as well as being the diagnostic criterion used to define Subunits ID and IF. Shore-based examination of smear slides has also identified a diatom-rich interval centered on ~118 mbsf (Fig. 3, Fig 4) that was initially missed and not given subunit status. Interestingly, all intervals rich in biogenic opal correspond to times of peak interglacial conditions (MIS 1, 5e, 9, and 11) immediately following abrupt glacial terminations. The implications of this observation are discussed below. Biogenic opal measurements that are not yet complete should soon provide a more rigorous and complete picture of opal deposition patterns in the Cariaco Basin over the past 580 ka.
Calcium carbonate and total organic carbon (TOC) data are presented in Figure 9. Measurements of carbonate and TOC were made at the University of British Columbia (by GHH) on samples spaced at ~30-cm intervals in Hole 1002C. The calcium carbonate content of the sediments was measured coulometrically, whereas percent TOC was derived from the difference between total carbon (analyzed using a Carlo-Erba CHN analyzer) and carbonate carbon. Analytical precision for the TOC and carbonate data is ±1.2 wt% and ±2 wt%, respectively.
The TOC and carbonate data from the Cariaco Basin show a strong relationship with the planktonic foraminiferal 18O record (Fig.
9). Although clear exceptions exist, sediments deposited during interglacial periods are generally characterized by both high TOC and carbonate contents. Interglacial TOC values range from averages of ~3 wt% in MIS 5, 7, 9, and 13, to as much as 5 wt% in MIS 1 and 11, whereas glacial TOC contents are as low as 0.1-1.3 wt%. In a similar fashion, the carbonate contents of interglacial sediments typically increase to between 20 and 40 wt%, whereas glacial carbonate values are generally less than 10 wt%. Superimposed on this glacial-interglacial pattern is evidence of higher frequency variability in both measured proxies; short-term TOC fluctuations, for example, are especially pronounced in MIS 3 sediments in a pattern similar to that observed in the record of 18O (Fig.
5).
The organic component of the Cariaco Basin sediments is thought to be predominantly of marine origin, although a minor terrigenous component may be present (Haug et al., 1998). Wakeham and Ertel (1988) reported 13O values of about -20 for the organic fraction of Holocene box core sediments, values typical of those observed in subtropical phytoplankton (Rau, 1994). In an analysis of eight depth-distributed samples from the old DSDP Site 147, McIver (1973) reported an average 13O of -21.0 for bulk organic matter, with values ranging from -20.4 to as light as -22.25 in one sample at 62 mbsf. McIver (1973) interpreted the latter value to reflect a modest increase in terrigenous contributions to the basin during a period of sea-level lowstand. To the extent that sub-bottom depths in Sites 1002 and 147 appear to be comparable (e.g.,
Fig. 2), this particular sample may have been deposited during MIS 6.
Coccoliths and foraminifers are the dominant source of biogenic carbonate in sediments of the Cariaco Basin, with small but significant contributions from pteropods (aragonite) over much of the sequence. Benthic foraminifers, ostracodes, and micromollusks in the >150-mm size fraction are present or largely absent as a function of the ventilation history of the basin, but probably do not contribute much to the total carbonate budget of the sediments. Although turbidite deposition appears to be minimal at the Site 1002 location on the Cariaco Basin's flat central saddle, the occasional presence of microturbidites (subcentimeter sized), molluscan debris, and abraded benthic foraminifers from the surrounding shelf in samples indicates the likely sporadic introduction of reworked materials into the sediment. The presence of reworked shelf carbonate may account for scattered high carbonate values (e.g., during MIS 8) that rise above the background pattern of carbonate variability (Fig. 9).
The abundance of terrigenous matter (weight percent) in the bulk sediment was calculated at a sampling interval of ~2 samples/1.5 m core section (Fig. 9) using a Ti-based normative calculation:
Concentrations of Ti were measured (by KMY and RWM) using ICP-emission spectrometry (ICP-ES) at Boston University, utilizing a suite of International Standard Reference Materials to assure precision (~2% of the measured value) and accuracy (within precision). This calculation assumes that terrigenous matter in the Cariaco Basin is similar chemically to Post-Archean Average Shale (PAAS) (Taylor and McLennan, 1985). While it is possible that this is not the case, PAAS is commonly used as a reference point for comparisons to other sediment. Furthermore, it has been shown that Ti concentrations in different "average shales" do not vary significantly (Murray and Leinen, 1993, 1996). Our normative calculation also assumes that all of the Ti in the sediment is mineralogically tied up in the terrigenous component. Though Ti is relatively particle reactive, variations in terrigenous matter using this equation cannot be attributed to adsorption by biogenic matter (e.g., Murray and Leinen, 1996) because such a biogenically scavenged component can only be detected in systems where biogenic matter comprises >95 wt% of the bulk sediment (Murray and Leinen, 1996). Titanium also does not respond to redox variations (Taylor and McLennan, 1985), so Ti variations in Cariaco Basin sediments are not diagenetically controlled. Thus, results shown in Figure 9 represent maximum estimates of the terrigenous contribution to each sample.
The Ti-based normative calculation indicates that the terrigenous matter content of Site 1002 sediments ranges from 36 to 91wt% of the total bulk composition, with an average contribution of 63 wt% ±11%. In general, the terrigenous contents of the sediments appear to have been high during glacial intervals and low during interglacials, a pattern perhaps not unexpected in a hemipelagic environment where climate and sea-level change are likely to have markedly affected sediment deposition.
![]()