RESULTS
Data for species common to the Antarctic and Subantarctic (e.g., Akiba, 1982; Johansen and Fryxell, 1985; Priddle and Fryxell, 1985; Fryxell et al., 1986; Baldauf and Barron, 1991; Fenner, 1991; Harwood and Maruyama, 1992; Hasle and Syvertsen, 1996) were combined to form an "Antarctic" (AA) group, representing diatoms transported from the Southern Ocean. The group comprises Asteromphalus hookeri, Asteromphalus parvulus, Azpeitia tabularis, Dactyliosolen antarcticus, Fragilariopsis kerguelensis, Proboscia barboi, Rhizosolenia praebergonii, Thalassiosira poroseriata, and Thalassiothrix antarctica. Of these, T. antarctica and P. barboi are the most abundant constituents of the AA group. Diatoms of the AA group were likely to have been living in Antarctic and Subantarctic open ocean waters. Those near the Antarctic polar frontal zone (APFZ) were likely to have been incorporated into the AAIW mass during its formation at this ocean front. Previous studies (e.g., Treppke et al., 1996) have also shown that displaced Southern Ocean diatoms present in the waters over the Walvis Ridge are transported within the AAIW. This watermass also serves as an important source of nutrients (Lange et al., 1999; Berger et al., 1998). Fluctuations in this group should, therefore, represent variation in the oceanography of the region, although changes in the productivity biomass cannot be ruled out. The DAR data for Chaetoceros resting spores and the AA group are shown in Figure F2.
The diatom assemblages at Site 1083 are dominated by Chaetoceros resting spores. Resting spore formation is associated with nutrient exhaustion in the water column, after mass multiplication of vegetative cells following an upwelling event (e.g., Bao et al., 1997, and references therein), often in highly-stratified waters (S. Boharty, pers. comm., 2001). Chaetoceros resting spores, therefore, provide a record of paleobloom events. Such qualities have led to their exploitation to great effect in many studies of both modern and paleo-upwelling regimes including southern Spain (e.g., Bárcena and Abrantes, 1998); northwest Iberia (e.g., Bao et al., 1997); California (e.g., Grimm et al., 1996; Hutchins and Bruland, 1998); Peru (e.g., Schrader and Sorknes, 1997), and the Antarctic region (e.g., Leventer, 1991; Rathburn et al., 1997). They have also been used in previous studies of the BC system (e.g., Treppke et al., 1996; Lange et al., 1999). Therefore, assuming that the abundance of Chaetoceros resting spores does not fluctuate in response to the changing intensity of biogenic silica dissolution, this proxy can be used with confidence to reconstruct productivity at Site 1083.
Chaetoceros resting spores comprise 33%-99% of the assemblage (averaging 76%), indicating that upwelling has been an important feature at this site throughout the whole of the time period investigated. This is in agreement with Lange et al. (1999), who reported their occurrence at Sites 1081, 1082, 1084, and 1085 in highly variable abundance throughout the same time period.
Prior to ~2.54 Ma, Chaetoceros resting spore abundance displays high sample-to-sample variability. Peaks in DAR occur with a periodicity of ~9-11 k.y. and maximum amplitude of ~4000 × 106 valves/cm2 per k.y. Maximum peaks in productivity occur during glacial Stages 104 (~3630 × 106 valves/cm2 per k.y. at ~2.62 Ma) and 102 (~3320 × 106 valves/cm2 per k.y. at ~2.57 Ma). Peaks of comparable magnitude also occur within interglacial Stages 105 (~3330 × 106 valves/cm2 per k.y. at ~2.649 Ma) and 103 (~2890 × 106 valves cm2 per k.y. at ~2.590 Ma). However, the latter peak is isolated so should be treated with caution. The data suggest overall that productivity was not significantly related to glacial/interglacial cyclicity at this time.
The DAR of the AA group is considerably higher and more variable between 2.65 and 2.54 Ma than the period following the INHG. Two sharp peaks in the DAR of the AA group correspond to those of Chaetoceros resting spores occurring at ~2.599 (~70 × 106 valves/cm2 per k.y.) and ~2.558 Ma (~55 × 106 valves/cm2 per k.y.).
After ~2.54 Ma, there appears to be a far more organized cyclicity within the Chaetoceros resting spores data. Peaks in the accumulation rate show an apparent cyclicity of ~30-35 k.y. Both the highest and lowest DAR values are found within this period during Stages 94 (~4250 × 106 valves/cm2 per k.y. at ~2.406 Ma) and 96 (~320 × 106 valves/cm2 per k.y. at ~2.447 Ma). As a consequence, the amplitude of the signal is much greater (an average of ~6400 × 106 valves/cm2 per k.y.). Other peaks of significantly elevated Chaetoceros resting spore productivity occur during glacial Stages 100 (~3800 × 106 valves/cm2 per k.y. at ~2.536 Ma), 98 (~4150 × 106 valves/cm2 per k.y. at ~2.497 Ma and ~3700 × 106 valves/cm2 per k.y. at ~2.492 Ma), and 96 (~4000 × 106 valves/cm2 per k.y. at ~2.457 Ma) and during the interglacial Stage 95 (~3395 × 106 valves/cm2 per k.y. at ~2.430 Ma). Peaks and troughs also appear to be more temporally sustained (i.e., a longer wavelength) where each oscillation encompasses a greater number of samples. This is especially so for the troughs found within the glacial Stages 100 and 96. Such temporal variability in DARs appears also to be a feature of the AA group, though on a comparatively lesser magnitude of scale.
The AA group DAR signal is suppressed following the INHG, but three prominent peaks occur at ~2.538 Ma (~80 × 106 valves/cm2 per k.y.), ~2.492 Ma (~75 × 106 valves/cm2 per k.y.), and ~2.428 Ma (~50 × 106 valves/cm2 per k.y.). The former two occur within glacial Stages 100 and 98, respectively, and the latter corresponds to a similar peak in Chaetoceros resting spores within interglacial Stage 95.
After ~2.54 Ma the major peaks in Chaetoceros resting spores occur at the interglacial-glacial transitions (i.e., during ice accumulation). DAR peaks rise and fall sharply across the boundaries. Such peaks tend to correspond with similar fluctuations in the AA group, suggestive of a relationship between the two.
Analysis of replicate laboratory standards produced error ranges of ± 0.2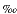 and ± 0.1 for
and ± 0.1 for  15N an 13Corg, respectively. Preliminary investigations of 13Corg values for Site 1083 (not shown here) range between -20.17 and -22.58, and average at -21.35 ± 0.1. This value indicates that the main source for the organic matter at Site 1083 is from marine algal production. This is in agreement with previous studies (e.g., by Treppke et al., 1996) that have shown the study area not to have been affected by input from the Cunene River. Therefore, the 15N signature of sediments from Site 1083 has the potential to contain a record of the nutrient utilization within the marine environment.
15N an 13Corg, respectively. Preliminary investigations of 13Corg values for Site 1083 (not shown here) range between -20.17 and -22.58, and average at -21.35 ± 0.1. This value indicates that the main source for the organic matter at Site 1083 is from marine algal production. This is in agreement with previous studies (e.g., by Treppke et al., 1996) that have shown the study area not to have been affected by input from the Cunene River. Therefore, the 15N signature of sediments from Site 1083 has the potential to contain a record of the nutrient utilization within the marine environment.
Where fluctuations in the 13Corg record should show changing productivity, this is related to the entire biological assemblage composing the organic matter. Fractionation may, therefore, be affected by changes within the entire biological assemblage of the site, and not necessarily reflect diatom abundance alone. For this reason, the 13Corg record is not discussed in detail here (see Maslin et al., unpubl. data).
The 15N values range between 6.11 and -0.45, and average 2.11 ± 0.2. The record shown in Figure F2 indicates great variability throughout the time period, but the overall relationship between 15N and Chaetoceros resting spores suggests NO3- limitation at times of low production. This is as expected since Chaetoceros resting spore formation is known to be sensitive to low NO3- conditions (e.g., Schuette and Schrader, 1979).
NO3- appears to have been at its least limiting during late Stage 101 and at the beginning of Stage 100 and most limiting during Stages 105 and 95. The difference between glacials and interglacials is not marked. However, there is an indication of reduced nitrate limitation during the interglacial-glacial transition periods following ~2.54 Ma.
This model of NO3- utilization remains tentative, however, because it assumes that the 15N signal is a reflection of NO3- utilization within the water column. In reality, the signal may be recording a response to a wide variety of factors, reflecting the complexity of the nitrogen cycle.
Fractionation of the isotopes may occur at any point within the cycle and both before and after assimilation by phytoplankton, therefore affecting the isotopic ratio in the environment. Key areas of alteration are highlighted in Table T1.
Of the potential sources of alteration, only terrigeneous input can be disregarded with confidence, because 13C results indicate that the sedimentary organic matter is marine in origin at Site 1083. However, in previous studies of the Benguela system (e.g., Holmes et al., 1998, 1997, 1996), only diagenesis and denitrification are consistently considered to affect the 15N record.
At present, the effects of diagenesis are poorly understood. For the Southern Ocean, Altabet and Francoise (1994) report 15N at the sediment/water interface was enriched by 5 to 9 compared to the surface waters. However, for the North Atlantic, Altabet et al. (1991) report a depletion of 15N with depth. Whereas the former is consistent with oxidative degradation of organic matter, the latter was attributed to a gain of 14N or a loss of 15N. Nevertheless, the majority of studies to date have shown that the 15N signal of the surface waters is transferred to the surface sediments, albeit with a constant offset. For the Walvis Ridge area, modern studies have shown this offset to be 1.6 (Holmes et al., 1998).
Denitrification, however, may be of importance, as it is likely to have some temporal variability. Most phytoplankton assemblages are dominated by cyanobacteria (Falkowski et al., 1998), so there is much potential for denitrification to take place, especially in zones of enhanced productivity. The modern Benguela system is known to be oxygen depleted, particularly near to the coast (Holmes et al., 1998). However, O2 is generally 1-2 mL O2/L, but denitrification takes place only when levels are <0.2 ml O2/L. Although concentrations this low are rarely observed on the shelf, they may occur. Studies of the modern Benguela system by Estrada and Marrasé (1987) showed that in April 1986, a subsurface NO3- minima coincided with a nitrite maxima and an oxygen minima, suggesting denitrification had taken place at 23°S. Boulégue and Denis (1983) also suggested that anoxia was responsible for both fish kills and enhanced sulfide production that have been observed on the shelf within the same area.
It is difficult to speculate on the effects of denitrification in this record. If such a process had taken place, it would manifest itself as a more positive excursion in 15N. This may have occurred at periods within the record when correlation with productivity indicators is poor (shown by asterisks in Fig. F2). The present data, however, is insufficient to indicate this with any great certainty. Nevertheless, productivity is still taking place at these points, suggesting that denitrification was not sustained and intense.
Similarly for the Chaetoceros resting spore record, TOC fluctuates with low amplitude within each climatic state throughout the time period prior to ~2.54 Ma, although levels are more elevated throughout Stage 104 (see Fig. F2). The lowest recorded TOC MAR occurs within this time period at ~2.593 Ma (21.06 g/cm per k.y.).
After ~2.54 Ma, the signals increase to more elevated levels, with records peaking at the interglacial-glacial transitions. This is particularly true for Stages 100 (~2.534 Ma) and 98 (~2.497 Ma), where there is a strong accordance with the record for Chaetoceros resting spores. However, where the Chaetoceros resting spores signal falls sharply after the initial peak during Stage 96, TOC MAR remains elevated.
Berger et al. (1998) suggested that silica might have been limiting within the Benguela region within the later stages of the Pleistocene. Silica (in the form of silicic acid) is a major requirement for diatoms for the construction of the frustule during cell division (e.g., Schrader and Schuette, 1981). Its limitation is less likely to affect nonsiliceous primary producers and may go some way to explaining the elevated TOC and chlorine MAR signals with a reduction in diatom production for this period. Alternatively, frustule dissolution in the water column is more likely to have taken place when concentrations of dissolved silica are reduced. Therefore, biosiliceous preservational issues must also be considered in this case.
