ORGANIC GEOCHEMISTRY
Headspace gas analysis was conducted as part of the standard protocol required for shipboard safety and pollution prevention monitoring. A total of 23 cores from Hole 1207A were evaluated (Table T10). The concentrations of CH4 were extremely low (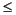 3 µL/L [ppmv]); no hydrocarbon gases higher than C1 were detected. The downhole profile of CH4 concentration in Hole 1207A shows a subsurface maximum in gas concentrations (Fig. F35), which is perhaps attributable to methanogenic bacterial activity in the near subsurface, albeit at levels barely above background. This interpretation is consistent with the occurrence of maximum sulfate concentrations at shallower depths (see "Alkalinity, Sulfate, Ammonium, Phosphate, Manganese, and
Iron" in "Inorganic Geochemistry"). Two samples from Hole 1207B were analyzed. The first was taken from the Campanian section (Core 198-1207B-6R), where the CH4 concentration matched those from comparable depths in Hole 1207A, and the second was taken from a disturbed Albian core (Core 198-1207B-28R). The highest CH4 concentrations for the site were observed in this latter sample, though they remain at trace levels (4.5 µL/L).
3 µL/L [ppmv]); no hydrocarbon gases higher than C1 were detected. The downhole profile of CH4 concentration in Hole 1207A shows a subsurface maximum in gas concentrations (Fig. F35), which is perhaps attributable to methanogenic bacterial activity in the near subsurface, albeit at levels barely above background. This interpretation is consistent with the occurrence of maximum sulfate concentrations at shallower depths (see "Alkalinity, Sulfate, Ammonium, Phosphate, Manganese, and
Iron" in "Inorganic Geochemistry"). Two samples from Hole 1207B were analyzed. The first was taken from the Campanian section (Core 198-1207B-6R), where the CH4 concentration matched those from comparable depths in Hole 1207A, and the second was taken from a disturbed Albian core (Core 198-1207B-28R). The highest CH4 concentrations for the site were observed in this latter sample, though they remain at trace levels (4.5 µL/L).
Carbonate and Organic Carbon
Carbonate determinations by coulometry were made for a total of 73 samples from Hole 1207A and Hole 1207B (Table T11). Samples were selected to provide a measure of the carbonate content within different units and to assess the influence of carbonate content on color reflectance. The values for carbonate range from 16 to 87 wt% (Table T11) within Subunit IA, reflecting variations in the proportions of siliceous and calcareous microfossils. The lowest value was an interval enriched in silica. The carbonate content profile shows alternating values downcore (Fig. F36). The variations observed are primarily an artifact of the dominant criterion for sample selection, namely the choice of intervals that are representative of extremes in color and lithology. However, some of the variations in these sediment properties likely are related to climatic fluctuations induced by orbital cycles. Carbonate contents are high (mean = 97.2 wt%) and show little variability in the Campanian calcareous oozes (Unit II).
Elemental concentrations of C, H, N, and S were measured (Table T12) for three samples from the carbonaceous interval correlated to the early Aptian OAE1a, hereafter described as upper, middle, and lower. Remarkably, the uppermost layer of the recovered interval of the black shale (upper; Sample 198-1207B-44R-1, 60-61 cm; 565.60 mbsf) contains 34.7 wt% organic carbon. A second sample from the central portion of the carbonaceous interval (middle; Sample 198-1207B-44R-1, 76-77 cm) contains 10.4 wt% organic carbon, and the base of the recovered interval (lower; Sample 198-1207B-44R-1, 103-104 cm) was determined to be 1.7 wt% organic carbon. However, these isolated values are indicators of the range of organic carbon values; they do not imply progressive enrichment through the section, especially given the inherent variability of Corg-rich horizons at other sites in the Pacific (Table T13). In addition, logging data from Site 1207 (Fig. F43) suggests that the actual thickness of the OAE1a horizon may be more than twice the interval recovered. The S content of the upper Corg-rich layer was also exceptionally high (10.6 wt%) (Table T12). Smear slide analysis shows that this sample contains abundant pyrite, although some of the sulfur exists in the form of organo-sulfur compounds (see below).
The C/N values for the lower, middle, and upper samples of the OAE1a are 16.2, 22.3, and 25.0, respectively (Table T12). In contemporary environments, high C/N ratios (>20-25) tend to reflect land-derived organic matter (Meyers, 1994), but Corg-rich horizons are exceptions to this tenet. Many mid-Cretaceous black shales (e.g., Meyers et al., 1984; Meyers, 1987) and Neogene or Quaternary sapropels (Meyers and Doose, 1999; Nijenhuis and de Lange, 2000) yield high C/N values that lie in the range typically indicative of terrestrial organic matter. Yet these units contain only minor contributions of material from land plants; visually, they are composed predominantly of amorphous organic matter derived from marine algae and bacteria. Consequently, the C/N ratios seem somewhat anomalous, which suggests that either the characteristics of the source organic matter are unusual and/or nitrogen cycling in these horizons may be radically different from the norm. Exceptional preservation of algal organic matter may reduce recycling of its nitrogen contents within the water column, thereby decreasing C/N ratios (Meyers and Doose, 1999). Perhaps the survival of cell walls that are rich in lipids and aliphatic biopolymers, like sheaths, significantly enhances Corg contents, without similarly increasing nitrogen content. C/N ratios would also be modified by preferential removal of nitrogen-rich organic compounds during anaerobic degradation of organic matter (Nijenhuis and de Lange, 2000). Moreover, nitrogen isotope profiles for Toarcian black shales provide evidence of partial denitrification in environments in which Corg-rich intervals are deposited (Jenkyns et al., 2001), and one might speculate that N-limitation led to higher C/N ratios. The recognition of high C/N ratios (31 to 590) in kerogens from the Archean and early Proterozoic (Beaumont and Robert, 1999) further confirms that the cycling of nitrogen behaves differently in anaerobic systems.
Rock-Eval investigations focused on samples from the early Aptian OAE1a to assess the origin of their kerogen. The hydrogen index (HI) and oxygen index (OI) values for the three samples generally are similar, despite the vast differences in their Corg contents (Table T13). The high HI and extremely low OI values confirm that Type I organic matter is dominant in all the samples (Fig. F37) (Tissot et al., 1974) and indicate that they are composed of organic matter of algal and bacterial origins. In addition, the relatively low Tmax values attest to the thermal immaturity of these samples.
Assessment of Extractable Hydrocarbons
The hydrocarbons in the lower, middle, and upper samples (198-1207B-44R-1, 103-104 cm; 44R-1, 76-77 cm; and 44R-1, 60-61 cm) from the lower Aptian OAE1a horizon were extracted to enable exploration of their biomarker compositions. The information obtained provides evidence of the biological sources and thermal maturity of the organic matter and of the likely depositional environment. The procedure followed the methodology described in "Organic Geochemistry" in the "Explanatory Notes" chapter. In addition, gas chromatography-mass spectrometry (GC-MS) was used to examine the biomarker composition of a CH2Cl2 eluate from the silica separation step of the middle sample consisting primarily of ketones.
The color of the extracts reflected their Corg contents. In particular, the color of the upper horizon extract was intense for such a small sample size. Also, during sample preparation by chromatographic separation, a pronounced pink coloration was evident in the polar (CH2Cl2) eluant, suggesting a significant porphyrin content. The hexane-soluble constituents, primarily consisting of aliphatic hydrocarbons, were analyzed by GC-MS and were identified from responses at specific target intervals in individual ion chromatograms and from their mass spectra. All of these components are well documented as components of mid-Cretaceous black shales (e.g., Barnes et al., 1979; Brassell et al., 1980, 1986; Rullkötter et al., 1984, 1987; Farrimond et al., 1986, 1990; Simoneit, 1986). Consideration of the principal characteristics of the samples (Table T14) and their relative abundances of biomarker components (Table T15) enables assessment and interpretation of aspects of the depositional environment of OAE1a.
Source of Organic Matter and Depositional Paleoenvironment
The extractable hydrocarbons are dominated by biomarkers of algal and bacterial origin, primarily short chain n-alkanes, and suites of polycyclic steroid and hopanoid hydrocarbons (Figs. F38, F39; Tables T15, T16). The abundance of n-C17 and the presence of branched alkanes, complemented by occurrences of methylhopanes, provide convincing evidence for contributions from cyanobacteria, especially in the lower sample. The steroids and hopanoids are derived from eukaryotic algae and bacteria, respectively (Table T15), although the absence of 4-methyl steroidal hydrocarbons suggests that dinoflagellates (Brassell and Eglinton, 1983; Robinson et al., 1984; Summons et al., 1987) were sparse among the plankton community. The only indication of allochthonous materials is the trace amounts of C25+ n-alkanes with pronounced odd/even preference that likely reflect inputs of vascular plant waxes borne by eolian dust (Simoneit, 1978; Zafiriou et al., 1985; Gagosian and Peltzer, 1986; Gagosian et al., 1987). The prevalence and character of bacterial components is consistent with the existence of a microbial mat, perhaps akin to those of the sulfur-oxidizing bacteria Thioploca in benthic settings (Williams and Reimers, 1983; McCaffrey et al., 1989; Ferdelman et al., 1997). Similarly, the organo-sulfur species, notably phytenyl thiophenes (Rullkötter et al., 1984; Brassell et al., 1986; ten Haven et al., 1990), confirm diagenetic incorporation of sulfur into organic moieties. The pristane/phytane ratios are less than unity (Table T14), which is typical of sediments associated with anaerobic systems (Didyk et al., 1978). In contrast, the presence of various ketones (Table T15) provides circumstantial evidence for oxidation of precursor lipids (Simoneit, 1973) or food web processes within the water column. However, steroidal ketones can also be biosynthetic products (Gagosian et al., 1982; Brassell and Eglinton, 1983; Robinson et al., 1984; Brassell et al., 1987) and consequently cannot provide conclusive, independent evidence of oxygenation levels in the water column or underlying sediments. In fact, the presence of lamination combined with the extremely high Corg contents is compelling evidence of anaerobic bottom water conditions.
Stratigraphic Variations in Biomarkers
The major variation in the biomarker signatures related to planktonic sources of organic matter is the evidence for more substantial cyanobacterial contributions in the lower sample. There are also subtle differences among the samples in their distribution of bacterial markers, reflecting either a hierarchical community or temporal changes in the planktonic or benthic bacterial populations. The hopanoid composition of the upper sample, combined with the extraordinary Corg and S preservation (Table T12), is consistent with the development of a microbial mat. By contrast, the steroidal distributions are remarkably uniform, suggesting little change in eukaryotic algal populations during OAE1a.
Summary
The catalog of biomarkers identified includes no significant constituents that do not originate from bacterial and/or algal sources (Table T15). Thus, the hydrocarbon composition of the intervals is consistent with the results from Rock-Eval pyrolysis that indicate algal and bacterial origins for the organic matter.
Organic-Rich Aptian Strata from the Mid-Pacific
Aptian organic-rich horizons have been recovered from several sites in the Pacific, including another site on Shatsky Rise (Site 305) (Table T13). Few detailed analyses of the organic geochemistry of these horizons have been performed. However, the availability of measurements of Corg contents combined with limited Rock-Eval data enables comparisons with results from shipboard analyses of the Aptian organic-rich interval from Site 1207. In the absence of carbon isotopic data, perhaps the best comparative measure is the modified van Krevelen diagram used to diagnose Rock-Eval data (Fig. F37), preferably complemented by biomarker compositions.
The Corg contents of Aptian samples from Site 1207 span the range associated with other organic-rich units. The high value of 34.7 wt% is comparable to that reported for the uppermost horizon of the expansive Aptian interval at Site 866, which contained 34.5 wt% Corg (Table T13). However, the organic-rich intervals at Site 866 show a disparate range of HI and OI indices that reflects the variable sources and/or degree of oxidation of organic matter in these shallow-water deposits. Several samples from Site 866 reveal affinities with terrestrial organic matter (i.e., OI > HI), whereas others appear to resemble the characteristics representative of algal kerogens, especially those from lacustrine systems. Moreover, inspection of published chromatograms (Baudin et al., 1995) reveals substantial differences between the aliphatic hydrocarbon distributions at Site 866 and Site 1207. It is difficult to properly compare the two sets of data in the absence of published identification of individual components within the chromatograms of Site 866 samples, although it is apparent that their n-alkane profiles are markedly different from those at Site 1207. Similarly, the comparative distributions of polycyclic aliphatic hydrocarbons, especially steranes and hopanes, appear to show significant disparities, based on simple efforts at peak identification by matching gas chromatography (GC) retention times.
Despite these discrepancies, there is one sample from Site 866 (143-866A-88R-1, 56-58 cm) (Table T13) that possesses Rock-Eval characteristics (Fig. F37) comparable to those from Site 1207 samples. Unfortunately, the absence of a published chromatogram for this sample precludes comparison of biomarker data. However, it lies within OAE1a recognized by the negative  13C excursion for carbonate that occurs immediately below Core 143-866A-188R (Jenkyns, 1995). Consequently, it suggests that there may be some uniformity in the organic matter associated with the early Aptian OAE1a event, which might imply conditions that exerted controls on the character of primary production and biomass.
13C excursion for carbonate that occurs immediately below Core 143-866A-188R (Jenkyns, 1995). Consequently, it suggests that there may be some uniformity in the organic matter associated with the early Aptian OAE1a event, which might imply conditions that exerted controls on the character of primary production and biomass.
A more extensive correlation between the HI and OI indices from the Site 1207 samples exists with samples from Site 463 that possess Corg contents exceeding 2.5 wt%. Other intervals at Site 463 with lower organic carbon contents tend to possess lower HI and higher OI indices. Poor resolution of individual components in the published chromatogram of the lower Aptian samples from this site (Samples 62-463-70-6, 29-35, and 39-42 cm) (Dean et al., 1981) hampers comparisons of biomarker compositions. Also, the assignments of C33 and C35 n-alkanes, based solely on GC retention time, appear erroneous. The same compounds are reported in Albian Sample 62-465A-38-1, 142-150 cm (Dean et al., 1981), but GC-MS analysis of a comparable Albian sample (from Section 62-465A-38-3) failed to detect them (Comet et al., 1981). Consequently, these peaks more likely correspond to hopanes, akin to those recognized at Site 1207. Reinterpretation of the identities of n-alkanes implies that these higher plant markers are absent and thereby eliminates the sole biomarker evidence for terrestrial organic matter in the lower Aptian at Site 463. Thus, algal/bacterial organic matter is dominant in the lower Aptian of both Sites 463 and 1207. Further comparison of their biomarker compositions is merited, and it seems pertinent to focus on Section 62-463-70-6 for two reasons. First, the Rock-Eval data for the Corg-rich sample from Sample 62-463-70-6, 39-42 cm (Table T13), are directly comparable to those from Site 1207, which place them adjacent on the van Krevelen diagram (Fig. F37). Second, this sample has a 13Corg of -27.7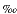 , which is the most negative value, albeit within a limited data set, from the lower Aptian at Site 463. Analysis of Sample 62-317A-16-2, 131-134 cm, from the Aptian at Site 317 presents a different picture. It has high Corg (24.7 wt%) (Table T13) and S contents (12.7 wt%), and the 13C value of its kerogen is -21.6 (Simoneit and Stuermer, 1982). Prominent among its biomarkers are the C31 n-alkane and 17b(H),21b(H)-homohopane, but it possesses a low relative abundance of pristine and phytane (PR/PH = 0.8) and no steroidal components. These biomarker characteristics, which have few parallels among the other lower Aptian samples, are indicative of deposition in a productive lagoon (Simoneit and Stuermer, 1982).
, which is the most negative value, albeit within a limited data set, from the lower Aptian at Site 463. Analysis of Sample 62-317A-16-2, 131-134 cm, from the Aptian at Site 317 presents a different picture. It has high Corg (24.7 wt%) (Table T13) and S contents (12.7 wt%), and the 13C value of its kerogen is -21.6 (Simoneit and Stuermer, 1982). Prominent among its biomarkers are the C31 n-alkane and 17b(H),21b(H)-homohopane, but it possesses a low relative abundance of pristine and phytane (PR/PH = 0.8) and no steroidal components. These biomarker characteristics, which have few parallels among the other lower Aptian samples, are indicative of deposition in a productive lagoon (Simoneit and Stuermer, 1982).
In summary, there is a consistency in Rock-Eval characteristics among specific intervals within the lower Aptian at Sites 463, 866, and 1207, whereas the biomarker characteristics for Site 317 appear anomalous. Further correlation among these sites awaits comprehensive evaluation by combining carbon isotopic profiles and detailed biomarker analysis in a high-resolution sample set.
Depositional Scenarios for OAE1a
The biomarker results have yielded perhaps the most informative indication of the depositional conditions of the early Aptian OAE1a event at Site 1207, namely evidence for cyanobacterial contributions from the occurrence of diagnostic branched alkanes in the lower sample. Recognition of the cyanobacterial contributions requires explanation. Are these diagnostic biomarkers observed because of enhanced preservation that effectively captured a molecular profile of the characteristics of oceanic phytoplankton typical of that time? Or does the evidence for cyanobacteria reflect their occurrence as biological harbingers of the conditions that triggered formation of the organic-rich intervals? Given the prominence of these biomarkers in the lower layer, it seems likely that the occurrence of cyanobacteria may be related to temporal variations in primary productivity associated with the early Aptian event. Perhaps ocean biomass and plankton populations changed in association with the advent of Corg-rich sediment deposition and then evolved further once environmental conditions conducive to the sequestration of organic matter were established and stabilized. A prominent role for cyanobacteria among plankton might suggest that nitrate was limiting. Evidence for water mass denitrification in the early Toarcian OAE (Jenkyns et al., 2001) demonstrates the critical controls on productivity exerted by nitrogen cycling and could help to explain the anomalously low C/N ratios for these Corg-rich intervals. Invocation of a phase of enhanced cyanobacterial production during OAE1a may also reflect a response to Fe availability that both triggers and stimulates increased phytoplankton production (Leckie et al., in press). Moreover, cyanobacterial lipids are depleted in 13C relative to their biomass (Sakata et al., 1997), and enhanced production and preservation of their hydrocarbon moieties therefore could potentially enhance the negative excursion in 13Corg associated with OAE1a (Menegatti et al., 1998; Bralower et al., 1999).
The subsequent changes in biomarkers that are manifested in the molecular stratigraphic records reflect evolving plankton communities, culminating in a population that seems to have included algae with colonial or mat affinities. Perhaps the last vestige of the early Aptian event sustained benthic microbial mats that enhanced the exceptional preservation of organic matter.
The extraordinary sequestration of organic matter (up to 34.7 wt%), even by comparison with other organic-rich intervals within the mid-Cretaceous, reflects a perturbation of the ocean-climate system that radically changed the processes controlling biogeochemical cycling. The profound effect on plankton productivity is recorded by exceptional preservation of organic matter from algal and bacterial sources, aided by anaerobic conditions. Progressive trends in phytoplankton populations survive in their molecular legacies, notably the indication of contributions from cyanobacteria within this OAE. Reconstruction of temporal changes in plankton communities and depositional conditions can be accomplished by elucidation of paleontological assemblages, isotopic C and N profiles and excursions, and evaluation of molecular signatures. These combined approaches offer complementary lines of evidence that can describe the biological response to this upheaval in carbon cycling and help define its cause.
