5. Site 12091
Shipboard Scientific Party2
PRINCIPAL RESULTS
Site 1209 is located in middle bathyal (2387 m) water depth close to the most elevated, central part of the Southern High of Shatsky Rise. The site is located on seismic line TN037-14A (Fig. F1). This profile is hard to correlate with other profiles on the southern and western flanks of the Southern High that are tied to drill holes. A tentative predrilling correlation with the Southern High seismic units of Sliter and Brown (1993) suggests a moderately thick Unit 1 (Neogene), an expanded Unit 2 (Paleogene), and a moderately thick Unit 3 (Upper Cretaceous). The site is close to the point where the stratigraphic sequence appears to be most complete; however, the section was expected to contain a number of minor disconformities as indicated by prominent horizontal reflectors. The total thickness of the sedimentary section at Site 1209 is estimated at ~1147 m. Basement underlying the site was formed during Magnetochron CM20 in the Tithonian (Nakanishi et al., 1989).
The major goals of Site 1209 drilling were to core a shallow, relatively expanded Paleogene and uppermost Cretaceous section. Holes 1209A and 1209B were cored largely with the advanced piston corer (APC). Only two cores were taken with the extended core barrel (XCB). Hole 1209C was drilled down to the lower Miocene, and then the Oligocene to Maastrichtian section was cored. The three holes terminated at different levels in the Maastrichtian due to difficulty penetrating chert horizons with the XCB center bit. The deepest recovered sediment was early Maastrichtian in age from 297.6 meters below seafloor (mbsf) in Hole 1209B.
Coring recovered a relatively thin (111.2 m) lower Miocene to Holocene (0-16.4 Ma) section of nannofossil ooze, clayey nannofossil ooze, and nannofossil ooze with clay (lithologic Unit I) (Fig. F2). This unit has an unconformity (11.19 to 15.1 Ma) separating the uppermost middle Miocene and the uppermost lower Miocene. Lithologic Unit I rests unconformably on lithologic Unit II (111.2 and 235 mbsf), which consists of nannofossil ooze and nannofossil ooze with clay of early Oligocene to early Paleocene age (28.6-65 Ma). Preliminary nannofossil and planktonic foraminiferal biostratigraphy suggests that the succession is largely complete. However, minor unconformities may be present, especially in the upper to middle Eocene interval (between 38 and 42 Ma) that is characterized by low sedimentation rates. Underlying lithologic Unit III of Maastrichtian age (65 to 70 Ma) is composed of nannofossil ooze and chert. Three prominent chert layers were encountered in the ~50-m Maastrichtian section in Hole 1209C. However, no horizons were found in the same interval in Hole 1209B, suggesting that chert layers are laterally discontinuous. The ooze surrounding the chert at the bottom of each hole was often still fluid and almost completely unlithified.
The upper middle Miocene to Holocene section at Site 1209 is apparently continuous and similar to contemporaneous sequences at Sites 1207 and 1208. Unconformities from the lower to middle Miocene and the lower Miocene to the lower Oligocene are partially equivalent to those observed at Sites 1207 and 1208, suggesting that regional oceanographic processes controlling erosion and dissolution had a major effect on sedimentation. The Neogene section at Site 1209 was deposited at much lower sedimentation rates than at the other sites, at least partially as a result of the lower production of biosiliceous and carbonate materials. Moreover, sedimentation at Site 1209 was exclusively pelagic, whereas the other two sites appear to have received a large supply of fine sediment from bottom water currents and eolian transport. A progression of decimeter- to meter-scale orbital cycles is observed in the sedimentary record at Site 1209. Preliminary ages suggest that dominant frequencies are eccentricity (100 k.y.) subsequent to 0.6 Ma, obliquity (40 k.y.) from 0.6 to 2.5 Ma, and then a gradual transition from long obliquity (1.25 m.y.) to long eccentricity (400 k.y.) to eccentricity (100 k.y.) through the lower Neogene and Paleogene section.
The lower Maastrichtian to lower Oligocene section at Site 1209 is also apparently complete. The highlights of Site 1209 coring were clearly associated with the recovery of the sedimentary record of several critical events in this interval, most of them in multiple holes. These include the Eocene-Oligocene transition, the Paleocene-Eocene Thermal Maximum (PETM), a prominent biological event in the early late Paleocene, the Cretaceous/Tertiary (K/T) boundary, and the mid-Maastrichtian deepwater event (MME). Sediments are almost completely unlithified, and the site appears to have remained above the calcite composite depth (CCD) for most of this time interval; hence, foraminiferal preservation is good to excellent. The stable isotope and paleontological records from Site 1209 will provide important information on the nature of environmental changes during the critical events and their effect on marine biotas. Site 1209 will provide a firm shallow end-member in the Shatsky Rise depth transect.
Eocene-Oligocene Transition
The Eocene to Oligocene transition represents the true end of the "greenhouse" world of the Mesozoic and Early Cenozoic. Although this transition occurred over a period of several million years, stable isotopic records reveal that much of the cooling occurred over a relatively brief (350 k.y.) interval in the earliest Oligocene known as Oi-1 (33.15-33.5 Ma) (e.g., Miller et al., 1991; Zachos et al., 1996). The deep oceans cooled by ~3°C during Oi-1, and large permanent ice-sheets became established in Antarctica (Zachos et al., 1992a, 1992b). Current reconstructions of ocean temperature and chemistry for the Eocene-Oligocene transition, however, are based primarily on pelagic sediments collected in the Atlantic and Indian Oceans (Miller et al., 1987; Zachos et al., 1996). Very few sections suitable for such work have been recovered from the Pacific (e.g., Miller and Thomas, 1985). As a consequence, we still lack a robust understanding of how global ocean chemistry or circulation evolved in response to high-latitude cooling and glaciation.
The Eocene/Oligocene boundary was recovered in Cores 198-1209A-14H, 198-1209B-14H and 15H, and 198-1209C-4H. Preliminary nannofossil and planktonic foraminiferal biostratigraphy suggests that the boundary interval is complete. The most marked feature in the transition record at Site 1209 is a gradual change, over ~7.5 m in the lowermost Oligocene and uppermost Eocene, from light brown to tan nannofossil ooze with clay to a light gray to white nannofossil ooze. A similar lithologic transition was observed in an identical stratigraphic position at Site 1208. However, the Site 1208 record is far more condensed than the Site 1209 record.
The distinctive color change in the Site 1209 record reflects a pronounced deepening in the CCD. This oceanwide event is thought to be related to the intensification of ocean circulation and/or to increased continental weathering (Zachos et al., 1996). Cycles from within the transition show that the transition was nonlinear and likely affected by orbital climatic variations. This observation will help constrain the cause of deepening of the CCD. Planktonic and benthic foraminiferal preservation in this interval is moderate. Thus, stable isotope stratigraphies from Site 1209 have the potential to provide a firm understanding of the evolution of Pacific surface and deep waters through this important climatic transition.
Paleocene-Eocene Thermal Maximum
At the other end of the climatic spectrum from the Eocene-Oligocene transition, the PETM is an abrupt and short-term (~210 k.y.) warming event at ~55.5 Ma that led to major transformation of marine plankton and benthos. This climatic event involved an increase of some 8°C of high-latitude surface water temperature and some 5°C of deep ocean water temperature. Warming is thought to have been driven by the input of a massive quantity of greenhouse gas, most likely methane, into the ocean-atmosphere system (e.g., Dickens et al., 1995). The response of the tropics and the Pacific Ocean to this climatic event is currently poorly known (Bralower et al., 1995).
The PETM at Site 1209 corresponds to a 12.5-cm-thick medium brown layer of clayey nannofossil ooze with a sharp basal contact and a gradational upper contact. This horizon was recovered in Sections 198-1209A-21H-7, 198-1209B-22H-1, and 198-1209C-11H-3. The detailed lithostratigraphy of the three PETM records recovered varies significantly on a millimeter scale as a result of deformation of the soft horizons during coring and splitting. Core 198-1209C-11H shows the most complete upper Paleocene to lower Eocene transition as indicated by multisensor track (MST) data. The event corresponds to a sharp change from a white nannofossil ooze to a brown nannofossil ooze with clay (Fig. F3). In Hole 1209B, these two lithologies are separated by an extremely thin (1 mm) dark brown clay seam. In the other two holes, this seam has been disturbed during coring and splitting.
The micropaleontological record of the PETM interval is summarized in Figure F3. Preliminary biostratigraphy shows that the event lies toward the top of nannofossil Zone CP8 and planktonic foraminiferal Zone P5. In addition, a single specimen of Gavelinella beccariiformis, a benthic foraminiferal species that goes extinct at the onset of the PETM (e.g., Thomas, 1990) was found in Section 198-1209A-21H-CC, several decimeters below the event (see "Biostratigraphy"). The abrupt decrease in the nannofossil Fasciculithus that occurs just above the PETM in other sections (Bralower et al., 1995, 1997; Aubry et al., 1996; Monechi et al., 2000) lies at the top of the clayey nannofossil ooze layer at Site 1209 (Fig. F3). The PETM layer corresponds to a prominent magnetic susceptibility peak followed by a 6- to 7-m interval of relatively high susceptibility that shows cyclic variation. The top of the event is currently undefined. However, preliminary biostratigraphy—in particular the first occurrence (FO) of the nannofossil Discoaster diastypus that lies 1.73 m above the base of the PETM (between Samples 198-1209A-21H-5, 130 cm, and 21H-6, 30 cm), and the last occurrence (LO) of the planktonic foraminifer Morozovella velascoensis that lies 2.38 m above the base of the event (between 198-1209A-21H-5, 129-130 cm, and 49-50 cm)—indicates that the section is condensed compared to continental margin records from the Atlantic and Tethys (i.e., Kennett and Stott, 1991; Norris and Röhl, 1999; Röhl et al., 2000).
The PETM interval is associated with dramatic turnover in nannofossil assemblages. One of the dominant nannolith genera, Fasciculithus, is replaced by Zygrhablithus bijugatus, a nannolith that is often a highly abundant or dominant component of Eocene assemblages (Fig. F3). The genus Discoaster is often highly abundant within the event itself, likely as a result of warming or increased oligotrophy (Bralower, in press). Common to abundant calcispheres are found in sediments from the PETM interval at Site 1209. These fossils are likely produced by calcareous dinoflagellates during intervals of adverse surface water conditions.
Low-latitude planktonic foraminiferal assemblages in the PETM also experienced a significant transformation with the sudden appearance of an ephemeral group of ecophenotpyes or new species of the genera Acarinina and Morozovella (Kelly et al., 1996). The range of these transient taxa is limited to the interval of the carbon isotope excursion; hence, they are known as the "excursion" taxa. Although preliminary observations did not yield any of the end-member excursion taxa, several forms that are intermediate between the true excursion taxa and their ancestors were observed. No detailed observations were made on benthic foraminifers at Site 1209 because of sampling limitations.
The response of the CCD is a sensitive indicator of change in carbon cycling during the PETM, likely as a result of the input of large quantities of methane into the ocean-atmosphere system (e.g., Dickens et al., 1997). This response can be monitored by changes in carbonate content and preservation. Nannofossil preservation below the event at Site 1209 is moderate, indicating that the site was located in the broad range of the lysocline. Preservation declines markedly in the 1-mm clay lens at the base of the event as shown by pervasive etching of coccolith shields and nannolith rims. Preservation remains poor for several centimeters above this level, but nannofossils never completely disappear from the record. This interval of poor preservation is accompanied by a high abundance of bladey rhombs that may be reprecipitated calcite. The deterioration in nannofossil preservation is evidence for an abrupt rise in the level of the CCD and lysocline during the PETM. The effects of this rise are not nearly as dramatic at Site 1209 as at Site 1208, which was situated almost 1 km deeper (see "Principal Results" in the "Site 1208" chapter).
Mid-Paleocene Biological Event
A prominent, 23-cm thick, dark brown nannofossil ooze with clay was found in Sections 198-1209A-23H-3, 198-1209B-23H-5, and 198-1209C-12H-CC. This layer shows a sharp magnetic susceptibility increase and a slight density decrease. Preliminary micropaleontological investigations suggest that this interval may represent a previously unrecognized event of considerable evolutionary significance. This interval lies within planktonic foraminiferal Zone P4 and coincides exactly with the evolutionary FO of the nannolith Heliolithus kleinpellii, an important component of late Paleocene assemblages and a marker for the base of Zone CP5. The interval immediately below the brown ooze layer contains significant numbers of the species Bomolithus elegans, which may be the ancestor of H. kleinpellii. The abundance of H. kleinpellii increases sharply in a transitional interval at the base of the brown ooze layer. Planktonic foraminifers in the ooze layer are characterized by a low-diversity, largely dissolved assemblage dominated by representatives of the genus Igorina. This low-diversity assemblage suggests some kind of oceanic perturbation.
A contemporaneous and prominent magnetic susceptibility peak was found on the Blake Nose at Site 1051 (U. Röhl et al.,
unpubl. data); this peak is associated with a dark brown chalk and correlates almost exactly with the FO of H. kleinpellii. Isotopic and further paleontological investigations are required to understand the oceanographic conditions that gave rise to this mid-Paleocene event as well as the detailed evolutionary relationships of the key taxa.
Cretaceous/Tertiary Boundary
At Site 1209 the base of lithologic Unit II coincides with the K/T boundary, marked by a significant lithologic change from uppermost Maastrichtian, white to very pale orange nannofossil ooze to basal Paleocene, darker grayish orange foraminiferal nannofossil ooze. This boundary is located in Sections 198-1209A-25H-6 and 198-1209C-15H-3 but occurs within an unrecovered interval in Hole 1209B. The most complete boundary sequence is found in Hole 1209C. Here the top of the Maastrichtian is slightly indurated, possibly indicating an incipient hardground, and overlain by lowermost Paleocene mottled, light orange, slightly indurated foraminiferal nannofossil ooze that grades into softer and paler tan-gray nannofossil ooze. The lowermost Paleocene layer is strongly bioturbated as shown by the pale orange roots within the irregular surface of the top of the white Maastrichtian ooze (Fig. F4). The basal Paleocene unit is ~14 cm thick and overlain by a 23-cm-thick pure white ooze. In Hole 1209A, the uppermost Maastrichtian white nannofossil ooze is separated from the lowermost Paleocene light orange foraminiferal nannofossil ooze by a watery, disturbed, orange brown clay horizon. This horizon was formed by disturbance during coring and splitting and does not correspond to the "clay horizon" that defines the K/T boundary in classic sections such as El Kef, Tunisia; Caravaca, southern Spain; and Gubbio (Bottaccione), Italy.
The micropaleontological record of the K/T interval is summarized in Figure F4. Preliminary biostratigraphy shows the well-known abrupt change in nannofossil and planktonic foraminiferal assemblages. The white nannofossil ooze (Hole 1209A: sample F2) yields a diverse, but dissolved, highly fragmented fauna of the latest Maastrichtian Abathomphalus mayaroensis planktonic foraminiferal Zone; the washed residue also contains rare, minute, well-preserved heterohelicids, predominantly Guembelitria, that suggests a possible preservation of Zone P0 fauna in the deepest burrows. Nannofossil assemblages below this interval (Hole 1209A: sample N1) are diverse, well preserved, and include relatively common Micula prinsii indicating correlation to the uppermost Maastrichtian nannofossil Zone CC26. The light orange foraminiferal nannofossil ooze burrows (Hole 1209C: sample F1) yield highly abundant, minute (<100 µm), and well-preserved planktonic foraminiferal assemblages that are dominated by heterohelicids and belong to the basal Paleocene Parvularugoglobigerina eugubina (P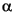 ) Zone. Nannofossils in the gray orange ooze (Hole 1209C: sample N2) are limited to "disaster" taxa (calcispheres) and reworked Cretaceous taxa. In the overlying pale nannofossil ooze horizon, the average size of the foraminiferal assemblage increases associated with increasing abundance of trochospiral forms with respect to heterohelicids (Hole 1209A: sample F3; Hole 1209C: sample F2). The lower part of the white ooze unit (Hole 1209C: sample N3; Hole 1209A: Sample N4) is dominated by ultrafine micrite, calcispheres, and the survivor coccolith taxon Cyclagelosphaera reinhardtii. Finally, the upper part of the white ooze unit (Hole 1209C: sample N4) contains fine micrite, C. reinhardtii, another survivor, Markalius inversus, and small species of the Danian coccolith genus Neobiscutum. This whole interval thus belongs to nannofossil Subzone CP1a. In Hole 1209A, the orange-brown soupy clay, containing a late Danian planktonic fauna, coats indurated ooze above the burrows fragmented by drilling.
) Zone. Nannofossils in the gray orange ooze (Hole 1209C: sample N2) are limited to "disaster" taxa (calcispheres) and reworked Cretaceous taxa. In the overlying pale nannofossil ooze horizon, the average size of the foraminiferal assemblage increases associated with increasing abundance of trochospiral forms with respect to heterohelicids (Hole 1209A: sample F3; Hole 1209C: sample F2). The lower part of the white ooze unit (Hole 1209C: sample N3; Hole 1209A: Sample N4) is dominated by ultrafine micrite, calcispheres, and the survivor coccolith taxon Cyclagelosphaera reinhardtii. Finally, the upper part of the white ooze unit (Hole 1209C: sample N4) contains fine micrite, C. reinhardtii, another survivor, Markalius inversus, and small species of the Danian coccolith genus Neobiscutum. This whole interval thus belongs to nannofossil Subzone CP1a. In Hole 1209A, the orange-brown soupy clay, containing a late Danian planktonic fauna, coats indurated ooze above the burrows fragmented by drilling.
The lowermost 2-3 cm of basal Paleocene darker grayish orange foraminiferal nannofossil ooze contains common spherules, probably altered tektites. The thin (1-2 cm) boundary clay unit that corresponds to planktonic foraminiferal Zone P0 in a few deep-sea and shelf sites (e.g., El Kef, Tunisia; and Caravaca, southern Spain) is not found at this site. However, Zone P0 fauna seems to be present in the deepest burrows.
We postulate that this interval has been affected by bioturbation soon after the boundary (Fig. F4). Nevertheless, the substantial thickness of the uppermost Maastrichtian M. prinsii (CC26) Zone and the lowermost Danian P Zone at Site 1209 indicates that the K/T boundary is paleontologically complete. In most deep sea-sites, the P Zone is either unrecovered or poorly preserved. Thus, the section represents one of the best preserved and least disrupted deep-sea records of this major extinction event, as well as of the subsequent radiation.
Mid-Maastrichtian Event
The long-term cooling of the Late Cretaceous was interrupted by a dramatic event in the mid-Maastrichtian (~69-70 Ma; nannofossil Zone CC24), when oceanic deep waters appear to have switched abruptly from low- to high-latitude sources (e.g., MacLeod and Huber, 1996; Frank and Arthur, 1999). This event appears to have coincided with the extinction of the inoceramid bivalves. Growing evidence, however, suggests that this biotic event is distinctly diachronous in the Atlantic, Tethys, and Pacific Oceans (MacLeod et al., 1996). Moreover, the magnitude and direction of stable isotope changes are quite variable from site to site (Frank and Arthur, 1999), possibly as a result of uncertainties in stratigraphic correlation or of true differences in deepwater properties.
The MME appears to have been recovered in Section 198-1209C-21H-1, which lies in nannofossil Zone CC24. In this section and several below it, large inoceramid prisms can be seen with the naked eye. Although the LO of inoceramids has not yet been determined in Hole 1209C, the abrupt disappearance is a likely sign of the event. Shore-based stable isotopic and benthic assemblage studies will help us refine our understanding of the origin and implications of this deepwater change.
1Examples of how to reference the whole or part of this volume can be found under "Citations" in the preliminary pages of the volume.
2Shipboard Scientific Party addresses can be found under "Shipboard Scientific Party" in the preliminary pages of the volume.
Ms 198IR-105
