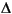 4-sterones relative to 5
4-sterones relative to 5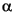 (H)-stanones in Samples 198-1213B-8R-1, 47-48 cm, and 63-64 cm (Table
T13), potentially reflect enhanced anaerobic sterol degradation observed in microbial mats (Edmunds et al., 1980).
(H)-stanones in Samples 198-1213B-8R-1, 47-48 cm, and 63-64 cm (Table
T13), potentially reflect enhanced anaerobic sterol degradation observed in microbial mats (Edmunds et al., 1980).
Headspace gas analysis was conducted as part of the standard protocol required for shipboard safety and pollution prevention monitoring. A total of five cores from Hole 1213A were evaluated (Table T9). The concentrations of CH4 were at background levels (range = 1.8-2.1 µL/L [ppmv]); no hydrocarbon gases higher than C1 were detected.
Carbonate determinations by coulometry were made for a total of 14 samples from Hole 1213A and 16 samples from Hole 1213B (Table T10; Fig. F30). Samples were selected to provide a measure of the carbonate content within different units.
The values for carbonate range from 11 to 80 wt% (Table T10) in Unit I, reflecting variations in the proportions of clay. The intervals with low values are condensed sections enriched in clay. Carbonate contents in the Cretaceous nannofossil chalks from Unit III range from 0.4 to 96 wt%. The lowest values correspond to lower Aptian organic-rich claystones deposited during OAE1a; variations derive from differences in clay and biogenic silica contents.
Elemental concentrations of C, H, N, and S were measured (Table T11) for nine samples from the lower Aptian to lower Valanginian (Subunits IIIC-IIIE) to assess the enrichment of organic matter within discrete intervals. In some samples, the darker horizons appear as stringers finely intercalated within carbonates; these prove to be lean in organic matter (0-0.4 wt%). The contents of organic carbon are highest in the lower Aptian samples. The two samples taken from the principal carbonaceous interval that correlates to OAE1a contain 25.2 and 10.2 wt% organic carbon. A third sample of clayey porcellanite interbedded with green claystone from lower in the section (Sample 198-1213B-8R-1, 96-97 cm) contains 2.9 wt% Corg. The organic carbon contents of Corg-rich intervals from the Valanginian were comparable (2.5 and 3.1 wt%). These horizons correspond to a minor peak in gamma ray logs (Fig. F39), whereas two more significant peaks are observed within the lower Aptian. The S contents of organic-rich layers range from 0.76 to 5.18 wt%; the highest value corresponds to the lower Valanginian horizon (Table T11). The C/N ratios of the organic-rich intervals range from 14.9 to 25.9 and are comparable to those reported for Site 1207, in part reflecting the different character of nitrogen cycling in these organic-rich intervals.
The focus for Rock-Eval investigations are samples from Corg-rich intervals, including the early OAE1a. The aim is to assess the origins of their kerogens, their thermal maturity and enable comparisons with Aptian Corg-rich intervals recovered at other sites.
The hydrogen index (HI) and oxygen index (OI) values for two of the lower Aptian samples (198-1213B-8R-1, 47-48 cm, and 63-64 cm) (Table T12) are similar despite the significant difference in their organic carbon contents (Table T11). High HI and extremely low OI values confirm that they are composed of type I organic matter (Fig. F31) (Tissot et al., 1974) derived from algal and bacterial sources. They are also similar to the lower Aptian samples from Site 1207 and the more Corg-rich intervals from Site 463 (Fig. F31). By contrast, the higher OI for the lower Aptian sample at 198-1213B-8R-1, 96-97 cm, which was interbedded with green claystone, is closer to type II kerogen. The high OI values for both Valanginian samples (198-1213B-15R-1, 9-10 cm, and 19R-1, 112-113 cm, respectively) (Table T12) can be attributed to the ineffectiveness of the Rock-Eval to fully exclude contributions from carbonate that enhance the S3 peak. This effect is well demonstrated by Sample 198-1213B-11R-1, 61-62 cm, which contained 65 wt% carbonate (Table T10). Consequently, the positions of these samples on the modified van Krevelen diagram are unrealistic. Unfortunately, the limited sample size precluded a second analysis after carbonate removal. The Tmax values, notably the exceptionally low value of 404°C for Sample 198-1213B-8R-1, 63-64 cm, attest to the thermal immaturity of these samples.
Hydrocarbon compositions were examined in samples (~1-4 g) from the lower Aptian (Samples 198-1213B-8R-1, 47-48, 63-64, and 96-97 cm), Valanginian (Samples 198-1213B-15R-1, 9-10 cm, and 19R-1, 112-113 cm) containing elevated Corg contents (>2 wt%) (Table T11). The information obtained provides evidence of the biological sources and thermal maturity of the organic matter and of the likely depositional environment. The procedure followed the methodology described in "Organic Geochemistry" in the "Explanatory Notes" chapter. In addition, gas chromatography-mass spectrometry (GC-MS) was used to examine the composition of CH2Cl2 eluates from the silica separation step of the samples consisting primarily of ketones.
The color of the extracts reflected their organic carbon contents. In particular, the color of the most Corg-rich horizons (Sample 198-1213B-8R-1, 63-64 cm) was intense for such a small sample size. During sample preparation by chromatographic separation, a pronounced pink coloration was also evident in the polar (CH2Cl2) eluants of Samples 198-1213B-15R-1, 9-10 cm, and 19R-1, 112-113 cm, suggesting that they contain high concentrations of porphyrins. The same eluate for another sample (198-1213B-8R-1, 96-97 cm) produced an intense yellow coloration, which could be associated with carotenoids, but their presence could not be confirmed by GC-MS. Both hexane and CH2Cl2 eluants, primarily consisting of aliphatic hydrocarbons and ketones, respectively, were analyzed by GC-MS, and their components were identified from responses at specific target intervals in individual ion chromatograms and from their mass spectra. All of the hydrocarbons are well documented as components of mid-Cretaceous black shales elsewhere (e.g., Barnes et al., 1979; Brassell, 1984; Rullkötter et al., 1984, 1987; Farrimond et al., 1986b, 1990; Simoneit, 1986). There are fewer published identifications of ketones in sedimentary rocks of this age (Comet et al., 1981; Farrimond et al., 1986a). Consideration of the principal characteristics of the samples (Table T13) and their relative abundances of biomarker components (Table T14) enables assessment and interpretation of aspects of the depositional environment of OAE1a and comparison with other intervals enriched in organic matter.
The extractable hydrocarbons are dominated by biomarkers of algal and bacterial origin, including short-chain n-alkanes and suites of polycyclic steroid and hopanoid hydrocarbons (Figs. F32, F33; Tables T15, T16). In the lower Aptian samples, the abundance of n-C17, complemented by occurrences of methylhopanes, provides evidence for contributions from cyanobacteria. The steroids and hopanoids are derived from eukaryotic algae and bacteria, respectively (see Table T15 in the "Site 1207" chapter), although the absence of 4-methyl steroidal hydrocarbons suggests that dinoflagellates (Brassell and Eglinton, 1983; Robinson et al., 1984; Summons et al., 1987) were sparse among the plankton community. The occurrence of alkenones in Sample 198-1213B-8R-1, 63-64 cm, further extends the temporal record of these markers for haptophyte algae (Farrimond et al., 1986a; Marlowe et al., 1990; Brassell, 1993). Their abundance, in concert with that of other functionalized lipids, strongly suggests that alkatrienones are unlikely to have been lost as a result of diagenetic alteration (cf. Farrimond et al., 1986a; Prahl et al., 1989; Hoefs et al., 1998). Thus, the absence of alkatrienones may reflect sea-surface temperatures that are elevated (i.e., >28°C) above the range where the composition of alkenones responds to temperature variations (Brassell et al., 1986; Prahl et al., 1988; Müller et al., 1998). Alternatively, this interval may precede the advent of temperature regulation of alkenone production and potentially the evolution of alkatrienone biosynthesis (S.C. Brassell, unpubl. data). The presence of sterol ethers in Sample 198-1213B-19R-1, 112 cm, is unprecedented in the Cretaceous (cf. Boon and de Leeuw, 1979; Brassell et al., 1980), although the origin of these components remains enigmatic (Brassell and Eglinton, 1986).
The only indication of allochthonous materials is the trace amounts of C25+ n-alkanes with pronounced odd/even preference that likely reflect input of vascular plant waxes borne by eolian dust (Simoneit, 1978; Zafiriou et al., 1985; Gagosian and Peltzer, 1986; Gagosian et al., 1987). The prevalence and character of bacterial components in the lower Aptian samples is again (cf. "Organic Geochemistry" in the "Site 1207" chapter) consistent with the existence of a microbial mat, perhaps akin to those of the sulfur-oxidizing bacteria Thioploca in benthic settings (Williams and Reimers, 1983; McCaffrey et al., 1989; Ferdelman et al., 1997). The pristane/phytane ratios are less than unity (Table T13), which is typical of sediments associated with oxygen-depleted systems (Didyk et al., 1978). In contrast, the occurrence of suites of steroidal ketones (Fig. F33; Table T13) provides circumstantial evidence for oxidation of precursor lipids (Simoneit, 1973) or food web processes within the water column. However, they can also be biosynthetic products (Gagosian et al., 1982; Brassell and Eglinton, 1983; Robinson et al., 1984; Brassell et al., 1987) and, consequently, cannot provide conclusive, independent evidence of oxygenation levels in the water column or underlying sediments. Certainly, the high total organic carbon (TOC) content and high HI of lower Aptian sedimentary rocks at Site 1213 suggest anoxic bottom-water conditions, but the lack of fine lamination and presence of burrows indicate at least periodic oxygenation at the sediment/water interface (see "Lithostratigraphy").
The catalog of biomarkers identified in the samples derive entirely from bacterial and/or algal sources (Table T14), consistent with the results from Rock-Eval pyrolysis.
The biomarker results from Site 1213 yield important information on the depositional conditions during early Aptian OAE1a. All of the Corg-rich intervals contain an abundance of steroidal ketones and suites of steroidal and hopanoid aliphatic hydrocarbons, which suggests a degree of uniformity in the contributions of organic matter, despite differences in organic carbon content. The steroids, in particular, reflect inputs from eukaryotes and are predominantly derived from planktonic algae, modified by water column predation. Steroidal ketones are abundant in the upwelling systems off Peru and Southwest Africa (Gagosian and Smith, 1979; Gagosian et al., 1983; Smith et al., 1983). There are also individual distinctive features, specifically the occurrence of alkenones and sterol ethers in Samples 198-1213B-8R-1, 96-97 cm, and 15R-1, 9-10 cm, respectively. Two specific features, however, distinguish the lower Aptian samples at Site 1213 and Site 1207 from the other organic-rich horizons, namely the presence of suites of methylhopanoids and extended hopanoids and the relative abundance of phytane and phytenyl lipids (phytenes at Site 1213 and phytenyl thiophenes at Site 1207). These characteristics can be attributed to contributions of organic matter from cyanobacteria and from methylotrophic and methanogenic bacteria. They imply the presence of specific bacterial communities associated with OAE1a, which in turn may reflect dysoxic conditions. However, the overall uniformity in the distributions of the dominant biomarkers, namely steroidal ketones, suggests that the populations of oceanic phytoplankton were stable over the time span represented by the samples. Yet, the higher prominence of 4-sterones relative to 5(H)-stanones in Samples 198-1213B-8R-1, 47-48 cm, and 63-64 cm (Table
T13), potentially reflect enhanced anaerobic sterol degradation observed in microbial mats (Edmunds et al., 1980).
The extraordinary sequestration of organic matter (>25 wt%) reflects a perturbation of the ocean-climate system that radically changed the processes controlling biogeochemical cycling. The potential roles of increased productivity and enhanced preservation in contributing to the survival of elevated amounts of organic matter are not easily distinguished. Yet, the majority of indicators for plankton productivity suggest only minor temporal variations in their populations, whereas the occurrence of components derived from bacteria suggests a change in the processing of organic matter at higher trophic levels. This evidence is most consistent with the concept that exceptional preservation of organic matter derived from algal and bacterial sources was aided by anoxic conditions.
![]()