BIOSTRATIGRAPHY
The three holes at Site 1259 recovered sediments of early Miocene to Cenomanian age. The lowermost cores yielded marginal marine to nonmarine sediments of presumable Cenomanian or Albian age. The marine succession contains calcareous nannofossils, planktonic foraminifers, and radiolarians in varying abundances and states of preservation, depending on lithology and sediment induration. The shipboard examination of these microfossil groups from core catcher samples was supplemented by additional samples from in the cores. The biostratigraphic data permitted zonal or stage assignments to be made for the entire sequence. Datum levels are summarized in Figure F11 and in Tables T3, T4, T5, T6, T7, T8, and T9.
A 26-m-thick nannofossil foraminiferal ooze (Cores 207-1259A-1R to 3R [upper part of lithostratigraphic Unit I]) at the top of the section has been attributed an early Oligocene age, reflecting a major slump or another type of redeposited sediment. This unit overlies a 342-m-thick, partly siliceous foraminiferal nannofossil ooze to chalk (Sample 207-1259A-4R-1, 0 cm, to 39R-5, 55 cm [lithostratigraphic Subunits IIA–IIC]) of early Miocene–earliest Eocene age, including the E/O boundary. Based on biostratigraphic data, two hiatuses were recognized in this interval: (1) the early Oligocene/early Miocene hiatus (Sections 207-1259A-10R-5 to 10R-6), with the late Oligocene being absent, and (2) the middle–late Eocene transition (Sections 207-1259A-15R-1 to 15R-2), with the proper boundary interval (planktonic foraminiferal Zone P15) missing. All microfossil groups studied are abundant and well preserved. An expanded section of 160 m of middle Eocene sediments was encountered.
The subsequent 75 m of clayey nannofossil chalk (Samples 207-1259A-39R-5, 55 cm, to 47R-3, 75 cm [lithostratigraphic Subunit IIIA]) is attributed to the Paleocene with a minor hiatus in its lower part, spanning parts of the early Paleocene (Core 207-1259A-46R). The P/E boundary is at 139 cm in Section 207-1259A-39R-5. At the base of the lowermost lowest Danian, a boundary clay and a spherule layer, which is believed to mark the base of the Tertiary, is developed. The K/T boundary sequence is considered to be complete or nearly complete because all the lower Danian and upper Maastrichtian biozones are represented in all holes.
The 50 m of upper Maastrichtian to mid-Campanian nannofossil chalk (Samples 207-1259A-47R-3, 75 cm, to 52R-5, 13 cm [lithostratigraphic Subunit IIIB]) is similar to that of the Paleocene, with calcareous microfossils being relatively rare and moderately preserved. A condensed glauconite-rich interval (80 cm thick) was recovered in Section 207-1259A-52R-4. It separates the Maastrichtian/upper Campanian chalk facies (lithostratigraphic Unit IIIB) from the ~56 m of black shales in lithostratigraphic Unit IV. Detailed high-resolution analyses of calcareous microfossils identify early Campanian assemblages at the top, early Campanian–Santonian assemblages in the upper third, early Santonian–late Coniacian taxa in the middle part, and early Coniacian taxa in the lowermost part of Section 207-1259A-52R-6. Cores 207-1259A-53R to 54R-CC are early Coniacian in age. Calcareous microfossils are generally present throughout the black shale. Foraminifers are often poorly preserved and of low diversity in calcareous interbeds, but pristine ("glassy") preservation is regularly found in clay-rich intervals. Calcareous nannofossils and planktonic foraminifers of early Turonian age are present near the base of the black shale. Section 207-1259C-18R-5 contains nannofossils of mid- to late Cenomanian age. A sharp contact between the laminated black shale and quartz sandstone (lithostratigraphic Unit V) was recovered in Core 207-1259A-60R. The latter unit, of which 1.40 m was recovered, did not provide any calcareous microfossils and was not dated. A more variable lithology (silty claystones and shelly sandstones) was encountered in the bottom part of Holes 1259B and 1259C.
As at the previous two sites, we recovered primarily Paleogene–mid-Cretaceous sediments from the three holes at Site 1259 that contain generally common to abundant calcareous nannofossils of moderate to good preservation. These allowed for zonal or stage assignments that are summarized in Figure F11 and Tables T3 and T7. Core catcher samples were examined for all holes and supplemented as necessary by samples in the cores for further refinement of the zonal assignments. These revealed major disconformities separating the following sedimentary packages:
- Approximately 28 m of a winnowed lower Oligocene foraminiferal sand and nannofossil ooze with some Miocene contaminants, apparently representing a slump deposit resting disconformably on the subjacent unit;
- Approximately 62 m of lower Miocene ooze of the same consistency, apparently in place, separated by another disconformity from the underlying unit;
- A ~34-m lower Oligocene interval separated by a disconformity from 6 m of upper Eocene calcareous ooze. This unit in turn is separated by a disconformity from the underlying 300-m middle Eocene–Paleocene nannofossil chalks. A minor disconformity may be present in the lowermost Paleocene;
- A 50-m Maastrichtian–late Campanian chalk beneath a complete K/T boundary; and
- An early Campanian–Cenomanian black shale sequence, unconformably overlying an undated quartz-cemented sandstone/gray clay sequence at the base.
In the more detailed descriptions that follow, the assemblages and ages pertain to Hole 1258A core catchers unless noted otherwise.
The first core catcher in the Oligocene slump or otherwise reworked interval contains abundant well-preserved Sphenolithus predistentus and Cyclicargolithus abisectus but no Sphenolithus distentus and is assigned to the lower Zone NP23 (Zone CP17); discoasters are overgrown. The subjacent Sample 207-1259A-2-CC yielded well-preserved Helicosphaera reticulata and common Reticulofenestra umbilica and Ericsonia formosa and is assigned to Zone NP21, assuming the latter two index taxa are not reworked. The third core catcher, however, is assigned to the younger Zone NP23, indicating an inverted sequence; it also contained a specimen of Coccolithus miopelagicus, which is of early Miocene age.
Samples 207-1259A-4R-CC and 5R-CC contain Sphenolithus belemnos and Triquetrorhabdulus carinatus along with abundant Hayaster perplexa, indicating a zonal assignment of uppermost NN2 Zone or, alternatively, NN3 if the T. carinatus is reworked (an assignment to Zone CN2 of the Bukry and Okada zonal scheme avoids that potential problem). Sample 207-1259A-6R-CC contains abundant H. perplexa and several large Discoaster druggii (up to 19 µm), the nominate species of Zone NN2. Although the sometimes elusive D. druggii was not noted in Samples 207-1259A-7R-CC and 8R-CC, the latter contains abundant and well-preserved T. carinatus, the acme of which is encountered near the base of Zone NN2, indicating that level or slightly lower. Sphenolithus disbelemnos is very abundant in Sample 207-1259A-9R-CC, suggesting an assignment of uppermost Zone NN1; Helicosphaera euphrates is abundant.
Abundant S. predistentus and C. abisectus along with common S. distentus are noted in the section in Sample 207-1259A-10R-CC, this time in normal stratigraphic order, and are assigned to the lower Oligocene Zone NP23 (upper part) (Zone CP18 of the Bukry and Okada compilation). A disconformity representing the missing upper Oligocene must lie within that core or in the core break above. The next three cores downhole (Cores 207-1259A-11R to 13R) contain S. predistentus in the absence of S. distentus, R. umbilica, and E. formosa, and belong to the lower portion of Zone NP23 (Zone CP17). Helicosphaera compacta is abundant.
The E/O boundary lies at a sharp color change (disconformity) in Section 207-1259A-14R-4, 27.5 cm. Lanternithus minutus is abundant above and Discoaster barbadiensis and D. saipanensis below the break. Specimens of these two discoasters are reworked immediately above the disconformity. Sample 207-1259A-14R-CC contains very large Cribrocentrum reticulatum (up to 12.5 µm), which assigns the sample to Zone NP20–19 (CP15). Cores 207-1259A-15R and 16R contain Dictyococcites bisectus and both probably belong to the upper part of Zone N17. Chiasmolithus grandis, however, was not observed until Sample 207-1259A-17R-CC, which, in the absence of D. bisectus, belongs to the lower part of Zone NP17, as does Sample 207-1259A-18R-CC.
A few Chiasmolithus solitus in the subjacent Sample 207-1259A-19R-CC might be reworked, but these are common in Cores 207-1259A-20R to 24R, along with R. umbilica, and are assigned to the lower part of Zone NP16 (or Subzone CP13B). Zone NP15 is assigned to Cores 207-1259A-26R to 29R based primarily on the presence of Chiasmolithus gigas, whereas Zone NP14 is assigned Sample 207-1259A-31R-CC on basis of a few specimens identified as heavily overgrown Discoaster sublodoensis.
Preservation and numbers of coccoliths in general continue to deteriorate downsection, making it difficult to distinguish a few possible Discoaster lodoensis in Core 207-1259A-32R, which also contains common Campylosphaera dela (some with crossbar intact), but no D. sublodoensis. This and the subjacent core are assigned to Zone NP13. Zeolites in the latter obscure the nannofossils, a condition not uncommon in this portion of the lower Eocene and where it is easiest to search for zonal markers using cross-polarized rather than plane-transmitted or phase-contrast light.
Common Tribrachiatus orthostylus, if not reworked in Sample 207-1259A-34R-CC, denote Zone NP12. A pristine specimen of Discoaster falcatus in the sample is reworked, however. Discoaster multiradiatus, D. diastypus, numerous "glassy" and sometimes misshapen rhombs, plus heavily overgrown triangular forms are interpreted as representing the Tribrachiatus bramlettei–Tribrachiatus contortus evolutionary transition; hence, this core catcher is assigned to Zone NP10.
Opal-CT lepispheres constitute the background of the partially silicified Samples 207-1259A-36R-CC and 37-CC, again making cross-polarized light the most practical way to search for coccoliths. Abundant Zygrhablithus bijugatus, D. multiradiatus, and somewhat misshapen "glassy" rhombs (interpreted as Rhomboaster spp.), however, indicate the upper part of Zone NP9. Neochiastozygus junctus is also abundant and continues, minus the rhombs but with occasional fasciculiths, down to the P/E boundary in Section 207-1259A-39R-3 or 39R-4. Below that level, fasciculiths replace N. junctus and Z. bijugatus in dominance and preservation improves considerably.
Sample 207-1259A-40R-CC yielded a variety of fasciculiths, D. lenticularis, and other characteristic Paleocene taxa such as Neochiastozygus concinnus. Zeolites are few in the subjacent clay-rich Sample 207-1259A-41R-CC, and preservation is quite good as indicated by pristine specimens of Toweius eminens; the sample belongs to the lower portion of Zone NP9. Helioliths are present in the subjacent core, and Discoaster mohleri persists downsection through an interval of nonrecovery (Core 207-1259A-42R) to Sample 207-1259A-43R-CC, which is assigned to the combined Zones NP7–NP8. The subjacent Sample 207-1259A-44R-CC contains common Ellipsolithus macellus, large Cruiciplacolithus tenuis (with well-developed "feet"), and common Lopholithus nascens but no helioliths. Below that, nannofossils are rare and poorly preserved in Sample 207-1259A-46R-CC, which yielded C. tenuis and possible Chiasmolithus danicus but no E. macellus; the sample was assigned to Zone NP3 of the Danian.
Immediately beneath the K/T boundary, Micula prinsii is present in Sample 207-1259A-47R-CC, which was assigned to Zone CC26. Its upper Maastrichtian ancestor, Micula murus, ranges downhole to Sample 207-1259A-49R-CC, which lies slightly below the lower CC26 zonal boundary. This species can be effectively substituted as the marker taxon for Zone CC26, as Nephrolithus frequens has hitherto not been observed in tropical latitudes.
In contrast to the expanded upper Maastrichtian section, the lower Maastrichtian is poorly represented if not absent entirely, as Core 207-1259A-50R is assigned to Zone CC25 and the subjacent core belongs to upper Zone CC23, based on the co-occurrence of Uniplanarius trifidum and Uniplanarius sissinghii, which top out at the Campanian/Maastrichtian boundary. This zone is traced downhole through Core 207-1259A-51R and into Sample 207-1259A-52R-3, 1 cm.
The transition into the black shales is well displayed in the photograph of Sections 207-1259B-18R-6 and 18R-7 (see "Lithostratigraphy") (Fig. F7). The normally greenish Campanian chalks have a dark gray color but with a greenish tinge at the base where glauconite appears. This glauconite-rich chalk continues downhole to the top of Section 207-1259B-18R-7, where Sample 18R-7, 20 cm, contains essentially the same upper Campanian nannofossil assemblage as above (U. trifidum, U. sissinghii, Micula decussata, and Aspidolithus parcus constrictus, etc., of Zone CC23).
Immediately beneath the green glauconitic chalk is a light yellowish to tan chalk separated from the chalk above by a heavily burrowed contact. Glauconite in this unit may have been introduced along with chalk from above by bioturbation; hence, care was taken to sample it where glauconite appeared to be absent. A "toothpick" sample 3 cm beneath the contact (Sample 207-1259B-18R-7, 27 cm) yielded common to abundant M. decussata, along with Eiffellithus eximius, A. parcus constrictus (two specimens), Broinsonia enormis, and Zeugrhabdotus bicrescenticus and a single but well-preserved specimen of the dissolution-resistant Marthasterites furcatus. This assemblage belongs to the lowermost Campanian Zone CC18, which is bounded at the top by M. furcatus and at the base by A. parcus.
The lithology changes rather abruptly again in Section 207-1259B-18R-7 from the light tan to a darkish brown claystone at ~53 cm, again with a heavily bioturbated contact. A coherent sample 3 cm below the contact at 56 cm also contained a sparse nannoflora with M. decussata, E. eximius, and B. enormis in the absence of A. parcus, the age of which could range from CC17 to CC14; planktonic foraminifers suggest a Santonian age, which would constrain the age of the sample to Zones CC17–CC15 (or CC18–CC15 if A. parcus is present). Several of the nannofossil datums traditionally used to further subdivide this interval are the dissolution-susceptible holococcoliths that are largely absent in the Cretaceous of the Leg 207 drill sites, such as Calculites obscurus (base of CC17) and Lucianorhabdus cayeuxii (base of CC16), hence the lack of precision in delineating these zones. We suspect that much of the Santonian is missing and that the lithologic change in Sample 207-1259C-18R-7, 53 cm, represents another hiatus.
Closely spaced samples beginning with Sample 207-1259A-52R-6, 2 cm, through 52R-6, 104 cm, recovered a condensed sequence of Santonian age. Sample 207-1259A-52R-6, 110 cm, continuing through 52R-6, 124–138 cm, contains E. eximius in the absence of both M. decussata and Lithastrinus septenarius and in the presence of occasional M. furcatus, all of which were assigned to the lower Coniacian Zone CC13. Marthasterites is common to abundant in the core catcher, and its acme seems to be characteristic of selected samples in the lower Coniacian of the Leg 207 sites. Cores 207-1259A-53R and 54R are also assigned to Zone CC13, with three samples (Samples 207-1259A-53R-1, 50 cm; 5R-CC; and 54-CC) also containing Quadrum gartneri. Nannoliths in the latter samples are common to abundant and well preserved.
Sample 207-1259A-55R-CC yielded no M. furcatus but rather the exclusively Turonian taxon Liliasterites angularis in the presence of E. eximius and Stoverius archylosus; Eprolithus eptapetalus is rare. The sample, therefore, is assigned to the mid- to upper Turonian Zone CC12.
Nannoliths are rare but well preserved in Core 207-1259A-57R, which is rich in zeolites and opal-CT lepispheres. Eiffellithus eximius is absent, but the numbers of E. eptapetalus are proportionately greater relative to the previous core catcher. This acme continues into the subjacent core catcher, and both cores are assigned to the lower Turonian Zone CC11.
Sample 207-1259A-59R-CC was problematic, as the very dark black zeolitic shale yielded rare and poorly preserved coccoliths consisting mostly of Eprolithus apertior, Eprolithus floralis, Zeugrhabdotus embergeri, Radiolithus planus, and small prediscosphaerids with small round bases ~6.5 µm across. An examination of samples in the core revealed L. angularis and Tranolithus orionatus in Sample 207-1259A-59R-1, 138 cm, and common Eprolithus octopetalus in Sample 59R-1, 138 cm, which together still indicate a Turonian age. A sample (207-1259C-18R-5, 75 cm), however, was dated as mid- to late Cenomanian in age based on the only specimen of Lithraphidites acutus observed on board ship during this leg.
Core 207-1259A-60R recovered only a silica-cemented quartz sand with cement particles that appeared opalescent in phase-contrast light. The two lowest core catchers in Hole 1259B (from Cores 207-1259B-25R and 26R) were both barren of nannofossils, the latter being a gray mudstone with numerous scattered 5- to 10-µm highly birefringed rhombs (dolomite?).
Planktonic foraminifer biostratigraphy at Site 1259 was based upon a combination of core catchers and samples taken from every section in Hole 1259A as well as from core catchers in Holes 1259B and 1259C. Zonal assignments are summarized in Figure F11 and Tables T4, T5, T8, and T9. Planktonic foraminifer Zones M2 (Miocene) through Zone KS20/KS22 (Turonian) were identified in Hole 1259A, along with significant breaks in the biozonation in the middle Oligocene–lowermost Miocene, the lower/middle Eocene boundary, and the upper Danian. Planktonic foraminifers were present in nearly all samples but varied widely in preservation and abundance. Preservation was best in clay-rich parts of the Turonian–Santonian sequence, although foraminifers were difficult to extract and clean in the black shales. Foraminifers in the light-colored bands of the black shales were frequently filled with calcite spar. Good preservation was found in the Miocene, Oligocene, and middle Eocene, whereas preservation was moderate or poor in the chalk of the lower Eocene, Paleocene, and Maastrichtian–Campanian sequences.
Sample 207-1259A-1R-1, 0–2 cm, contains a mixture of modern diatoms together with modern planktonic foraminifers such as Orbulina universa, Globigerinoides sacculifer, Neogloboquadrina dutertrei, and Globigerinoides ruber. The fine fraction (<150 µm) includes fragments of Oligocene foraminifers and pieces of manganese crust. Pleistocene sediments are only a few centimeters thick and were nearly entirely washed off the core top in Hole 1259A, whereas the core top was relatively well preserved in Hole 1259B.
Yellow calcareous ooze in Sample 207-1259A-1R-CC contains a diverse lower Oligocene foraminifer assemblage including common Turborotalia ampliapertura, Turborotalia euapertura, Globoquadrina galavisi, Chiloguembelina cubensis, and Cassigerinella chipolensis, representing Zone P19. In contrast, Sample 207-1259B-1R-CC contains markers for Oligocene Subzones P21b and P21a and Zones P20 and P19, together with Pleistocene foraminifers. Likewise, Samples 207-1259A-2R-CC and 3R-CC are also of Oligocene age but decrease in age downhole from Zone P18 to Subzone P21a, suggesting either massive contamination of the core catchers in the soupy foraminifer sands or massive reworking. Extensive reexamination of Sample 207-1259A-1R-CC did not reveal any reworked components.
The surficial Oligocene foraminifer sands are likely to represent a slump or reworking, since Samples 207-1259A-4R-CC to 9R-CC contain a succession of foraminifers representing lower Miocene foraminifer zones in stratigraphic order. Samples 207-1259A-4R-CC to 7R-2, 50–54 cm, contain a typical Zone M2 fauna with Globoquadrina dehiscens, Globigerinoides primordius, Globoquadrina binaensis, Dentoglobigerina altispira, and Globigerinoides altiapertura. The species Globorotalia mayeri, Globigerinella obesa, Globigerina angulosuturalis, and Globoturborotalita woodi are secondary components. A small four-chambered, heavily encrusted foraminifer resembling Neogloboquadrina pachyderma was found in most Miocene samples and has been provisionally called Neogloboquadrina nympha after the species described originally by Jenkins (1966) from the Southern Ocean.
Sample 207-1259A-7R-3, 50–54 cm, contains the highest record of Fohsella kugleri in Hole 1259A and marks the top of Subzone M1b. The base of this zone is marked by the first appearance of G. dehiscens in Sample 207-1259A-9R-7, 50–54 cm. Typical assemblages in Zone M1 include G. mayeri, F. kugleri, Globoquadrina venezuelana, Globoquadrina praedehiscens, and G. altiapertura. Subzone M1a, defined by the presence of F. kugleri in the absence of G. dehiscens, extends to Sample 207-1259A-10R-5, 50–54 cm.
A significant unconformity representing the upper Oligocene and possibly part of the basal Miocene occurs between Samples 207-1259A-10R-5, 50–54 cm, and 10R-6, 50–54 cm. Overlying sediments are assigned to Zone M1a based on the presence of the zone marker species F. kugleri, whereas sediments below the boundary contain abundant C. cubensis, G. angulosuturalis, and C. chipolensis, indicating lower Oligocene Zone P21a. Between Samples 207-1259A-11R-2, 50–54 cm, and 11R-3, 50–54 cm, the disappearance of G. angulosuturalis and the absence of Turborotalia ampilapertura suggests Zone P20. However, Sample 207-1259A-11R-5, 50–54 cm, contains G. angulosuturalis, suggesting that the sample is either contaminated with material reworked downhole or that the overlying sediments are incorrectly dated as Zone P21a. Zone P19 occurs between Samples 207-1259A-11R-CC and 13R-3, 50–54 cm, and is characterized by the occurrence of Turborotalia ampilapertura, C. chipolensis, C. cubensis, Globoquadrina tripartita, and Globoquadrina globularis. Finally, Zone P18, characterized by Pseudohastigerina spp., was found between Samples 207-1259A-13R-4, 50–54 cm, and 14R-3, 50–54 cm.
We have not observed large-scale inversions of zonal assignments as seen in the Oligocene at Site 1260. However, there are age reversals that raise a flag of caution about the continuity of the section. For example, G. angulosuturalis, the marker for Subzone P21a, is present in a few samples from both Zones P18 and P19, including Sample 207-1259A-13R-CC, where it is common. These occasional apparent age reversals at Site 1259 could be the result of downhole contamination, but at least some of the reversals occur in chalks that are not notably disturbed by rotary coring. The other possibility is that the whole section was extensively reworked during the late early Oligocene or perhaps prior to the deposition of Miocene sediments.
The contact between the Eocene and Oligocene occurs in Sample 207-1259A-14R-4, 28 cm. Zone P18 assemblages are juxtaposed with those of Zone P16 that contain Turborotalia cunialensis, Hantkenina alabamensis, Turborotalia pomeroli, and Globoquadrina pseudovenezuelana. In turn, Zone P16 assemblages are in contact with Zone P14 faunas between Samples 207-1259A-15R-1, 50–54 cm, and 15R-2, 50–54 cm. Typical species in Zone P14 include Turborotalia cerroazulensis, Morozovella spinulosa, Morozovella lehneri, Planorotalites renzi, H. alabamensis, and Acarinina rohri. Zone P14 can be traced as low as the interval between Samples 207-1259A-19R-2, 50–53 cm, and 19R-3, 50–53 cm.
The remainder of the middle Eocene is well represented in Hole 1259A and consists of an expanded and nearly continuous sequence of biozones and datum levels. Zone P13 is present between Samples 207-1259A-19R-3, 50–53 cm, and 20R-4, 50–54 cm. These samples show the evolution of Orbulinoides beckmanni from a taxon with three chambers in the last whorl that is similar to large Globigerinatheka kugleri or Globigerinatheka euganea to a fully spherical variety in which the final chamber constitutes half or more of the total shell. The lower limits of Zone P13 are defined by the first appearance of large globigerinathekids that resemble the holotype of O. beckmanni, a specimen that is similar to a large example of G. kugleri (see discussion of Zone P13 in
"Calcareous Nannofossils" in the "Site 1260 chapter"). Common species in Zone P13 are Igorina broedermanni, M. spinulosa, T. pomeroli, M. lehneri, and Acarinina bullbrooki. Zone P12 ranges from Sample 207-1259A-20R-5, 50–53 cm, to 23R-CC, whereas Zone P11, whose top is defined by the last appearance of Morozovella aragonensis, extends between Samples 207-1259A-24R-1, 50–54 cm, and 27R-1, 50–54 cm. Morozovella lehneri and M. aragonensis are both present between Samples 207-1259A-24R-1, 50–54 cm, and 24R-CC. Finally, Zone P10 was identified between Samples 207-1259A-27R-2, 50–54 cm, and 30R-CC, with its top defined by the first appearance of G. kugleri and its base approximated by the first appearance of Guembelitroides nuttali. Typical species in Zone P10 include M. aragonensis, Acarinina praetopilensis, Muricoglobigerina senni, Acarinina punctocarinata, A. bullbrooki, and P. renzi.
The lower Eocene may contain hiatuses in Zones P9 and P7 because these zones are relatively thin at Site 1259. The lower boundary of Zone P9 is located between Samples 207-1259A-33R-1, 50–54 cm, and 33R-2, 50–54 cm, based upon the lowest occurrence of fragmentary remains of Planorotalites palmerae. Assemblages in Zone P9 contain large specimens of Acarinina aspensis, with six or more chambers in the last whorl, in addition to A. praetopilensis, Pseudohastigerina micra, A. bullbrooki, I. broedermanni, and M. aragonensis. Zone P8 is represented by an assemblage that includes large "Globigerina" lozanoi, Acarinina pentacamerata, M. aragonensis, Acarinina quetra, and P. renzi in the absence of both P. palmerae and Morozovella formosa. Samples 207-1259A-33R-6, 50–54 cm, to 33R-CC are assigned to Zone P8. Zone P7 is recognized by the presence of M. formosa together with M. aragonensis and was observed only in Sample 207-1259C-1R-CC. In as much as Zone P7 represents ~1.5 m.y. of geologic time, it is likely that there are either unrecognized hiatuses in the biozone or a highly condensed section. In contrast, Zone P6 is represented by an expanded section between Samples 207-1259A-34R-CC and 38R-4, 52–55 cm. The foraminifer fauna consists of Acarinina soldadoensis, Morozovella subbotinae, M. aequa, Morozovella gracilis, and Acarinina wilcoxensis (major elements) and Pseudohastigerina sp., M. formosa, Morozovella lensiformis, and Morozovella marginodentata (secondary elements). We did not subdivide Zone P6 using the first occurrence of M. formosa because of very poor preservation in Samples 207-1259A-35R-CC to 37R-CC.
The P/E boundary occurs in Cores 207-1259A-39R, 207-1259B-7R, and 207-1259C-7R based upon a clay-rich horizon and the appearance of a distinctive fauna of "excursion taxa" (Kelley et al., 1998) as well as Chiloguembelina wilcoxensis, Pseudohastigerina sp., and I. broedermanni. The top of Zone P5 is indicated by the extinction of Morozovella velascoensis and occurs between Samples 207-1259A-38R-3, 52–55 cm, and 38R-4, 49–52 cm. A core gap in Hole 1259A prevents the detailed identification of the base of Zone P5, which occurs somewhere between Samples 207-1259A-41R-CC and 43R-CC.
A set of four small (~5 cm3) samples were taken in the ~80-cm interval above the base of the P/E boundary interval in Section 207-1259C-7R-2 to document the preservation and species composition of the foraminifer assemblage associated with the boundary. The base of the distinctive green clay horizon occurs at 120 cm in this section, and Samples 207-1259C-7R-2, 100–102 cm, 7R-2 108–110 cm, and 7R-2 115–117 cm, were barren of planktonic foraminifers, although all contained small numbers of benthic foraminifers and abundant but poorly preserved spumullarian radiolarians. Sample 207-1259C-7R-2, 70–72 cm, contains a moderately well preserved foraminifer assemblage with abundant Morozovella allisonensis and A. soldadoensis and rare Acarinina africana, Acarinina sibaiyaensis, Subbotina patagonica, and Parasubbotina varianta. Three of these species, M. allisonensis, A. africana, and A. sibaiyaensis, are associated with P/E boundary sections in other tropical and subtropical sites (Central Pacific, Egypt, Spain, New Jersey, and the Blake Plateau) and are known as the excursion fauna because of their abundance and near restriction to the Paleocene/Eocene Thermal Maximum (PETM) (Kelly et al., 1998). An unusual element of these assemblages is Parasubbotina paleocenica, a clavate species that has previously been reported only from ODP Site 1220 in the equatorial Pacific and its type area in coastal Senegal. The West African and equatorial Pacific settings of these previous discoveries suggest that P. paleocenica is associated with upwelling conditions. Hence, we infer that the excursion fauna may also represent an expansion of relatively productive waters during the PETM.
All the Paleocene zones are represented at Site 1259. Zone P4 is present between Samples 207-1259A-43R-CC and 45R-1, 50–54 cm, based upon the presence of Globanomalina pseudomenardii, Acarinina nitida, and Acarinina subspherica, all of which make their appearance in this zone. Subzone P3b occurs between Samples 207-1259A-45R-2, 50–54 cm, and 45R-CC. Typical species in Subzone P3b include M. velascoensis, Morozovella angulata, Morozovella pasionensis, Subbotina velascoensis, Subbotina triangularis, and Igorina albeari—a fauna similar to that of Zone P4 but lacking G. pseudomenardii and the acarininids. We have not found markers for Subzone P3a in any hole at Site 1259, but Zone P2 is well represented between Samples 207-1257A-46R-CC and the top of Core 207-1259A-47R by a low diversity assemblage of Praemurica inconstans, Praemurica uncinata, Morozovella praeangulata, Parasubbotina pseudobulloides, and Globanomalina compressa. A sample per section in Core 207-1259B-12R reveals a Subzone P1b fauna in Sample 207-1259B-12R-2, 94–95 cm, with Subbotina triloculinoides, Praemurica pseudoinconstans, and P. pseudobulloides, a Subzone P1a assemblage in Sample 207-1259B-12R-3, 67–68 cm, with Praemurica taurica and Eoglobigerina eobulloides, and a P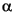 fauna with Parvularugoglobigerina eugubina, Guembelitria cretacea, and Woodringina sp. in Samples 207-1259B-12R-4, 53–54 cm, 12R-CC, and 13R-1, 24–25 cm. The spherule bed that is believed to mark the base of the Danian is in Section 207-1259B-13R-1, 49 cm.
fauna with Parvularugoglobigerina eugubina, Guembelitria cretacea, and Woodringina sp. in Samples 207-1259B-12R-4, 53–54 cm, 12R-CC, and 13R-1, 24–25 cm. The spherule bed that is believed to mark the base of the Danian is in Section 207-1259B-13R-1, 49 cm.
The Maastrichtian occurs between Core 207-1259A-47R and 50R. Sample 207-1259C-8R-7, 66–68 cm, taken ~16 cm below the spherule bed, yielded a diverse assemblage of uppermost Maastrichtian foraminifers including Abathomphalus mayaroensis (marker species for Zone KS31) and Plummerita hantkenoides, a species known to appear shortly before the Cretaceous/Tertiary (K/T) boundary. At best, P. hantkenoides is a rare species in most open ocean sites but its presence in Site 1259 suggests the upper Masstrichtian preserves an unusually complete record of latest Cretaceous paleoclimate and evolution. The combination of cyclostratigraphy and magnetostratigraphy that seems likely to be available at Leg 207 sites should allow the range of P. hantkenoides to be determined precisely during postcruise studies. As at other Leg 207 sites, Zone KS31 is represented by rare to common Rugoglobigerina rugosa, Globotruncanita stuarti, Pseudoguembelina costulata, Pseudotextularia elegans, Rugoglobigerina rotundata, Rosita contusa, Gansserina gansseri, and Globotruncana aegyptiaca. Occasional specimens of Globotruncanita calcarata and Globotruncanita subspinosa are found in most Maastrichtian samples, suggesting regular reworking from the Campanian. Zone KS31 is present between the K/T boundary and Sample 207-1259A-48R-7, 50–54 cm. The presence of R. contusa in the absence of A. mayaroensis suggests that Subzone KS30a (the R. contusa–R. fructicosa Zone) is present in Samples 207-1259A-48R-CC to 49R-CC. The absence of R. contusa in Samples 207-1259A-50R-1, 50–54 cm, to 50R-CC that still contain G. gansseri suggests Subzone KS30b, which spans the Campanian/Maastrichtian boundary.
Campanian foraminifer assemblages are poorly preserved and represented by only rare individuals. The G. aegyptiaca Zone (KS29) is represented in Sample 207-1259A-51R-CC by Rugotruncana subcircumnodifer, Globotruncana lineana, Globotruncanella petaloidea, and G. aegyptiaca along with an abundance of fish bones, glauconite pellets, and benthic foraminifers. Assemblages that may represent either Zone KS29 or KS28 are found in Samples 207-1259A-52R-3, 49–52 cm, and 52R-4, 123–126 cm, in a glauconite-rich horizon that overlies organic-rich claystones at the top of the black shale sequence. These samples contain a diverse but flattened foraminifer fauna with Heterohelix globulosa, Globotruncanella havanensis, R. rugosa, R. subcircumnodifer, Gansserina wiedenmayeri, Globotruncanita stuartiformis, and Globotruncanita esnehensis. Sample 207-1259A-52R-5, 26–28 cm, contains mostly benthic foraminifers and pyrite nodules but also Globigerinelloides prairiehillensis and R. rugosa, suggesting a Campanian age.
In contrast to the Campanian foraminifer assemblages in the glauconite-rich horizon, the underlying brown organic-rich claystones in Sample 207-1259A-52R-6, 111–113 cm, contain a distinctive and well-preserved Santonian assemblage with Marginotruncana coronata, Dicarinella concavata, Rosita fornicata, Archaeoglobigerina blowi, and Dicarinella canaliculata that in aggregate suggest Zone KS24. Coniacian foraminifers are found in Samples 207-1259A-52R-CC to 54R-CC and include both spar-filled and glassy empty-chambered foraminifers representing Zone KS23. Species in the Coniacian sequence include Marginotruncana sinuosa, Marginotruncana schneegansi, Dicarinella imbricata, Dicarinella primitiva, and Whiteinella inornata.
Turonian foraminifers were found starting in Sample 207-1259A-55R-1, 130–132 cm, to the lowest identifiable foraminifer assemblages in Sample 59R-CC. Species diversity is relatively high in the upper half of this sequence but decreases to only one or two species that are not diagnostic of either Turonian or Cenomanian age. The upper part of the Turonian includes an abundance of whiteinellids, such as W. inornata with weak imperforate bands, Whiteinella archaeocretacea, and Whiteinella baltica, along with the nearly ever-present Hedbergella delrioensis. Dicarinella imbricata and D. canaliculata are also present in many samples. The clavate species, Hastigerinelloides watersi, Hastigerinelloides amabilis, and Hastigerinelloides alexanderi, are rare components. There is low species diversity in foraminifer assemblages from Sample 207-1259A-58R-4, 42–45 cm, to 59R-CC and consists of little more than H. globulosa, Heterohelix moremani, H. delrioensis, W. archaeocretacea, and W. baltica, which could indicate either a Turonian (KS20/KS21) or latest Cenomanian (KS20) age. Foraminifers are often small (<180 µm) but are frequently preserved as glassy specimens with empty chambers in the clay-rich parts of the sequence. Foraminifers are entirely absent from Sample 207-1259A-59R-CC and are represented only by a few specimens of H. moremani in Sample 207-1259A-59R-CC, along with rare very well rounded quartz grains. The underlying quartz sandstone and claystone sequence in Samples 207-1259B-24R-CC and 25R-CC did not yield any foraminifers.
Radiolarians at Site 1259 were found in most of the cored material but were abundant and well preserved only in the middle Eocene. Core catcher samples were examined systematically from Hole 1259A, with an additional limited number of samples taken from Holes 1258B and 1258C.
The earliest Miocene Zone RN1 was identified in Samples 207-1259A-8R-CC and 9R-CC, based on the marker species for the base of the zone (Cyrtocapsella tetrapera) and Calocycletta serrata, which last occurred at the upper part of Zone RN1.
In the middle Eocene interval, the abundant occurrence of species Podocyrtis goetheana in Samples 207-1259A-17R-CC and 18R-CC marks the base of Zone RP16. This combination with the presence of species Spongatractus pachystylus (last occurrence [LO] at the top of Zone RP16) in Sample 207-1259A-16R-CC allows restriction of the age of Cores 207-1259A-18R through 16R solely to Zone RP16. Podocyrtis chalara is very abundant in Samples 207-1259A-19R-CC and 20R-CC, whereas Podocyrtis mitra is absent in the former sample but present, although very rare, in the latter. These samples are, therefore, situated above the base of the evolutionary transition from P. mitra to P. chalara (which defines the base of Zone RP15). Sethochytris triconiscus and abundant P. mitra in Sample 207-1259A-21R-CC allows its assignment to the upper part of Zone RP14. Therefore, the boundary between Zones RP15 and RP14 is placed in Core 207-1259A-21R. Podocyrtis fasciolata and Podocyrtis trachodes were observed in Sample 207-1259A-22R-CC, which can be therefore assigned to the lower part of Zone RP14 or the top of Zone RP13. Samples 207-1259A-23R-CC and 24R-CC are assigned to the upper part of Zone RP13 based on the occurrence of abundant Podocyrtis ampla, Podocyrtis sinuosa, and Eusyringium fistuligerum.
Sample 207-1259A-25R-CC contains both Podocyrtis phyxis and P. ampla, but the former is more abundant than the latter. It is, therefore, placed below the base of Zone RP13. As rare representatives of species E. fistuligerum are present in this sample, its age can be restricted to the top of Zone RP12. Therefore, the boundary between Zone RP12 and RP13 is placed in Core 207-1259A-25R.
Cores 207-1259A-25R down to 29R are assigned to Zone RP12 based on the presence of species Thyrsocyrtis triacantha. Sample 207-1259A-29R-CC contains both T. triacantha and its ancestor, Thyrsocyrtis tensa. As the former is more abundant than the latter, Sample 207-1259A-29R-CC can still be assigned to Zone RP12. Radiolarian preservation is moderate to poor in Sample 207-1259A-30R-CC and deteriorates further downhole. The co-occurrence of Dictyoprora mongolfieri and Thyrsocyrtis robusta suggests that it can be assigned to the lower part of Zone RP12 or RP11.
With respect to the Upper Cretaceous and, more particularly, the Campanian, Sample 207-1259A-51R-CC contains abundant radiolarian specimens (i.e., Amphipyndax sp. and Dictyomitra sp.) of moderate preservation that could not be identified at the species level. Sample 207-1259A-57R-CC contains moderately well preserved but rare radiolarians. The presence of Acanthocircus hueyi allows its assignment to the Alievium superbum Zone (lower Turonian or younger).
