RESULTS
Diatom data from 38 samples are shown on Table T2 and Figure F3. Diatoms are well preserved and abundant in most samples from Hole 1098C. Preservation of diatoms is excellent in the laminated sediments, suggesting burial by rapid deposition and little postdepositional destruction. Diatom preservation is diminished in the bioturbated intervals of Hole 1098C. Abundance of diatoms, expressed as millions of valves per gram of sediment, is high throughout the hole but is consistently high in interval C (Table T2). Highest diatom abundance is also observed in a comparable depth interval in Hole 1098B (Sjunneskog and Taylor, in press; Taylor and Sjunneskog, in press).
Four diatom taxa (Chaetoceros resting spores, Fragilariopsis kerguelensis, Thalassiosira antarctica, and the Fragilariopsis curta/Fragilariopsis cylindrus group) comprise the majority of the species in this study. Most of the other taxa make up a minor component of the assemblages, although in laminated sediments, nearly monospecific assemblages of Corethron criophilum, Rhizosolenia spp., Proboscia spp., and Thalassiothrix sp. are commonly observed. An exception is that the Fragilariopsis ritscheri/Fragilariopsis obliquecostata group is more abundant in interval B and Fragilariopsis angulata is more common in interval D (Fig. F3).
The relatively small number of samples examined for this study make the recognition of patterns and associations between the diatom assemblages and changes in other proxies within individual intervals difficult. Another problem is the species-specific composition of many of the samples that may represent a single diatom bloom event (e.g., Sample 178-1098C-1H-6, 10-12 cm). However, several changes between the intervals are evident. For example, in intervals A and B, F. kerguelensis is consistently less abundant than the F. curta/F. cylindrus group, but in intervals C and D, F. kerguelensis is more abundant than the F. curta/F. cylindrus group.
An interesting characteristic of interval D is the increased abundance of the asymmetric form of Eucampia antarctica. This trend has also been recognized from comparable depths in Hole 1098B (16.79-29.58 mbsf) (Sjunneskog and Taylor, in press). The symmetric forms of E. antarctica are believed to be a more "polar" form, whereas the asymmetric form is identified as a "more northerly" form (Fryxell, 1989; Kaczmarska et al., 1993). Table T3 shows the high ratio of symmetric/asymmetric forms of the species E. antarctica in two samples from interval D (average = 0.645). This ratio can be compared with the modern sediments of the Palmer Deep (core LGM-98-02 KC1, 1-71 cm), where this ratio has an average value of 3 (J. Murray, pers. comm., 2000).
Introduction
As can be seen in Table T1, benthic foraminifers occur in all samples from Hole 1098C. Preservation of the agglutinated foraminifers ranges from excellent to poor. The robust Milliamina arenacea and the abundant watch glass-shaped Deuterammina glabra are present in every sample (Fig. F4), whereas the organically cemented Portotrochammina eltaninae and poorly cemented Textularia spp. occur more sporadically and are often very fragile. Percentage values for 24 of the most commonly occurring species are presented in Tables T4, T5, T6, and T7. Rarely occurring benthic species are combined as "other agglutinated" or "other calcareous" species (see "Appendix
A", for species names). Calcareous foraminifers are poorly preserved in Hole 1098C and are often frosted, pitted, and partially dissolved. B. aculeata is the most common calcareous species, followed by Bolivinella pseudopunctata (Tables T4, T5, T6, T7). The planktonic foraminifer Neogloboquadrina pachyderma sinstral occurs rarely in Hole 1098C (Table T8).
Whereas calcareous foraminifers vary from common to absent in Hole 1098C, their interpretation is greatly complicated by post-core recovery dissolution. In May 1999, ~14 mo postcruise, several levels of Hole 1098C were resampled to obtain additional calcareous foraminifers for 14C dating. We found horizons that yielded common calcareous foraminifers in the original August 1998 samples were essentially barren of calcareous foraminifers in May 1999. Table T9 shows comparison of resampled horizons. For the remainder of the discussion, we must assume that the record of calcareous foraminifers from Hole 1098C has been degraded by an unknown but substantial amount of carbonate dissolution. Downcore changes in the abundance and composition of calcareous foraminifer species may reflect either original variations in the total assemblage, the influence of post-recovery dissolution, or any possible combination of these factors. In addition, we also recognize that there may have been postdepositional dissolution at this site that cannot be documented. Our results can only document an almost total dissolution of the calcareous fauna between August 1998 and May 1999. The timing of dissolution that occurred prior to August 1998 is unknown.
A characteristic feature of Palmer Deep benthic foraminifers that was recognized early in our study is the small size of the agglutinated fauna (Table T10). In most samples, the majority of the calcareous species are found in the larger sand fraction (>100 µm), whereas the majority of the agglutinated species are small and occur most abundantly in the smaller sand fraction (63-100 µm). We suspect the size distribution of calcareous foraminifers is due to the selective dissolution of the smaller calcareous specimens prior to the August 1998 sampling. Summary statistics and averages of agglutinated and calcareous foraminifers in Hole 1098C are located in Table T1. Because of the unknown amount of calcareous faunal dissolution, agglutinated benthic (ABF) and calcareous (CF) foraminifers per gram were calculated separately. In addition, the benthic foraminifer accumulation rate (BFAR) (Herguera and Berger, 1991) was calculated with calcareous foraminifers only, as is usual, and was also calculated as the agglutinated benthic foraminifer accumulation rate (ABFAR) (Table T1). There is no evidence for the selective destruction of agglutinated tests within Hole 1098C. In fact, the large numbers of very poorly cemented agglutinated foraminifers indicate that lab processing is not an explanation for foraminiferal loss.
Results of Foraminifer Analysis
The abundance and diversity of foraminifers fluctuate in Hole 1098C (Tables T1, T4, T5, T6, T7). Interval B contains the highest numbers of calcareous foraminifers and benthic foraminiferal species. Interval D contains the second largest numbers of calcareous species (Table T1) and also the highest number of planktonic foraminifers, which occur in approximately half of the samples (Table T8). Interval D is also different from the other intervals studied in that both the calcareous benthic foraminifer B. pseudopunctata and the agglutinated P. eltaninae are rare to absent (Table T7). As a result, D. glabra reaches its maximum abundance in interval D (Fig. F4; Table T7). Interval D also contains the largest percentages of rarely occurring calcareous species (defined as the total of all calcareous species except B. aculeata and B. pseudopunctata) (Table T7). Interval C contains the highest percentage of agglutinated foraminifers and the highest accumulation rate of agglutinated foraminifers (ABFAR on Table T1). Samples from interval C contain the fewest average number of calcareous foraminifers (Table T1), but isolated samples contain common calcareous foraminifers (up to 75% of the total foraminiferal fauna in one sample) (Table T6).
The R-mode cluster analysis of the 24 benthic agglutinated and calcareous foraminifer species resulted in three clusters that described the two end-member associations (laminated vs. bioturbated). Cluster 1 contains D. glabra and M. arenacea, cluster 2 contains P. eltaninae and Textularia weisneri, and cluster 3 consists of all the calcareous species with one rare agglutinated species (Fig. F5).
A Q-mode cluster analysis of the 125 samples identified core intervals that were dominated by the D. glabra and M. arenacea assemblage (cluster 1), the P. eltaninae and T. weisneri assemblage (cluster 2), and a calcareous assemblage (cluster 3). Samples characterized by cluster 3 included all samples where calcareous foraminifers were abundant (>30% of the total fauna) (Fig. F6). Samples characterized by clusters 1 and 2 (agglutinated species) are samples with low numbers of calcareous foraminifers and lower MS. In addition, interval D does not contain any samples of cluster 2 (P. eltaninae).
In each of the four intervals studied, there is a correspondence between laminations, MS, and the type of benthic foraminifers present (Fig. F6). Generally, laminated sediments with relatively low MS are dominated by agglutinated species (clusters 1 and 2) and massive sediments with fluctuating but relatively high MS are characterized by the same agglutinated species along with calcareous foraminifers (cluster 3) (Figs. F4, F6). We suggest that this pattern reflects syndepositional conditions that may have been modified by postdepositional dissolution. This pattern is most pronounced in intervals A, B, and D and is weak but present in the strongly laminated interval C.
Comparison to Core PD92-30
In an effort to quantify the amount of calcareous foraminiferal dissolution in Hole 1098C, a comparison was done with the original Palmer Deep piston core PD92-30. Core PD92-30 was shipped whole to the Antarctic Research Facility at Florida State University, where it was split and sampled ~6 mo after collection. Because there was no time between the splitting and sampling of core PD92-30, a comparison between the faunal counts of core PD92-30 and Hole 1098C could help estimate the carbonate loss that occurred during the 6 mo between the splitting and sampling of Hole 1098C.
Using the MS records, we could easily identify intervals of core PD92-30 that were comparable to intervals A and B in Hole 1098C. Table T11 compares the number of calcareous and agglutinated benthic foraminifers in each sample and per gram in the comparable intervals of core PD92-30 and Hole 1098C. It can be seen that the numbers of calcareous foraminifers in each sample and per gram are quite similar in both Hole 1098C and core PD92-30. The slightly higher values for the number of calcareous foraminifers in Hole 1098C probably result from the unequal number of samples used in the comparison. The significant differences in the number of agglutinated foraminifers probably results from their very small size (Table T10), which may have caused them to be uncounted in the previous study (Leventer et al., 1996).
The comparable numbers of calcareous foraminifers in both core PD92-30 and Hole 1098C suggest that both cores were probably affected by calcareous dissolution prior to sampling. However, the reduced time between splitting and sampling of core PD92-30 suggests that dissolution was not exacerbated during the expanded time interval between splitting (March 1998) and sampling (August 1998) of the Hole 1098C samples. All of this points to dissolution being both a syndepositional and postdepositional process in the Palmer Deep Basin and possibly other high-latitude sedimentary basins.
The calcium carbonate compensation depth (CCD) was approximated at 900 mwd along the AP (Ishman and Domack, 1994), which may help to explain carbonate loss in the Palmer Deep. Nevertheless, calcareous dissolution has continued during storage of Hole 1098C core. Likewise, samples from core PD92-30 collected in August 1999 have confirmed that dissolution of calcareous foraminifers has also occurred in the core repository (Table T12).
Agglutinated Foraminifers
Because of the dissolution of calcareous foraminifers in Hole 1098C, any valid paleoceanographic interpretation must be made using the abundant agglutinated assemblage. Unfortunately, agglutinated foraminifers are understudied. Because of their rarity (0%-16% of an assemblage) and poor downcore preservation, agglutinated foraminifers are usually ignored in paleoceanographic studies in favor of the more abundant calcareous species (i.e., Mead and Kennett, 1987; Ishman and Domack, 1994; Ohkushi et al., 2000). Today, agglutinated assemblages are extremely rare and restricted to the deep ocean below the CCD (Gooday, 1990; Evans and Kaminski, 1998; Osterman et al., 1999) or tidal marshes (Goldstein and Watkins, 1990; Scott et al., 1990). To test the value of agglutinated species in paleoceanographic studies, Murray and Alve (1994) subjected calcareous assemblages from the North Atlantic shelf, slope, and abyss to dissolution by acetic acid. The acid-treated samples resulted in a high diversity agglutinated assemblage that showed distribution patterns that correlated with recognized North Atlantic water masses. Murray and Alve (1994) concluded that high diversity agglutinated assemblages of the fossil record were the result of partial or total loss of the calcareous element through either syn- or postdepositional dissolution. Furthermore, the resulting agglutinated assemblages were as useful and perhaps even more useful than calcareous foraminifers for paleoceanographic analysis (Alve and Murray, 1995; Murray and Alve, 1999).
Support for the use of agglutinated foraminifers in Antarctic paleoceanographic studies can be found in several studies. The CDW-influenced diatom oozes of Marguerite Bay contain a diverse assemblage composed of 77% agglutinated foraminifers, and three agglutinated species comprise 14%-50% of the total assemblage in the eight CDW samples but rarely occur elsewhere along the AP shelf (Ishman and Domack, 1994). Harloff and Mackensen (1997) also identify an agglutinated assemblage that characterizes the CDW and the most organic carbon-rich sediments of the Scotia Sea above the CCD. Whereas this fragile agglutinated assemblage would not likely be preserved and would be represented in the geological record of the Scotia Sea as barren intervals (Harloff and Mackensen, 1997), the preservation of agglutinated species is not a major problem in the geologically young and rapidly deposited Palmer Deep sediments. Therefore, several studies have reported a dominantly agglutinated assemblage in association with diatom oozes of the CDW.
Discussion of Foraminiferal Results
In general, the foraminifer accumulation rates (ABFAR and BFAR) can be used as a measure of surface-water productivity (Herguera and Berger, 1991; Herguera, 2000). The high BFAR and ABFAR in Hole 1098C (Table T1) attest to the high biogenic productivity of the Palmer Deep (Table T2). However, the decay of organic matter in highly productive basins often results in bottom waters reduced both in oxygen and pH. Syndepositional carbonate dissolution caused by lowered pH is one explanation for dominantly agglutinated benthic foraminiferal assemblages found in Hole 1098C. This process may also explain the dominantly agglutinated foraminiferal assemblage from the surface diatom ooze at ODP Site 741, Prydz Bay, Antarctica (561 mbsf) (Schröder-Adams, 1990), and the CDW-influenced Marguerite Bay (Ishman and Domack, 1994) and the Scotia Sea (Harloff and Mackensen, 1997).
Rapidly-accumulating, laminated, organic-rich sediments are also associated with anoxic basins, but the benthic foraminifers of Hole 1098C do not support this conclusion. Agglutinated benthic foraminifer morphologies have been shown to be useful in the identification of bottom-water oxygen levels (Kaminski et al., 1995; Nagy et al., 1995). The two most common agglutinated species of the Palmer Deep, the watch glass-shaped D. glabra and troccaminid P. eltaninae, are both classified as mobile epifaunal species that are not tolerant of dysoxic conditions (Kaminski et al., 1995). Low-oxygen but not dysoxic conditions are also suggested by the patchy distribution of benthic foraminifer species (Tables T4, T5, T6, T7) (Douglas, 1987) and the high numbers of foraminifers (Table T1) that often result from the absence of macrobenthos competition (Bernhard and Reimers, 1991).
Carbon and oxygen isotopic values were measured in 75 samples (see "Appendix
B"). The sample data are restricted primarily to the massive high-susceptibility sediments where calcium carbonate is preserved. Only the shallow infaunal species B. aculeata was available for isotopic analyses in our samples. This taxon is not generally used in isotopic studies, but there was no alternative species available. Results of  18O are shown on Figure F6. The sporadic occurrence of calcareous foraminifers, the infaunal nature of B. aculeata, and the unknown effect of carbonate dissolution hamper interpretation of isotope data from our samples. We prefer to not overinterpret the data.
18O are shown on Figure F6. The sporadic occurrence of calcareous foraminifers, the infaunal nature of B. aculeata, and the unknown effect of carbonate dissolution hamper interpretation of isotope data from our samples. We prefer to not overinterpret the data.
In general, we see little variation in the isotopic data (see "Appendix
B"). There is some evidence that suggests that B. aculeata may calcify different chambers at depth within the sediment. This habit would result in each test providing an average isotope value over various sediment slices (Mackensen et al., 2000). This seems to be supported by our research, and oxygen isotope values only vary by ~0.2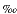 within each sample interval. Values in intervals A, B, and C range between +3.6 and +3.8 (Fig. F6). Interval D shows similar variation (0.2) in isotope data with the exception that oxygen isotope values are slightly more positive, varying between +3.8 and +4.0.
within each sample interval. Values in intervals A, B, and C range between +3.6 and +3.8 (Fig. F6). Interval D shows similar variation (0.2) in isotope data with the exception that oxygen isotope values are slightly more positive, varying between +3.8 and +4.0.
Carbon isotope values show slightly more variability, although most analyses range between +0.0 and -0.5. Values in interval D are on the average slightly lower than those in intervals A, B, and C. We see no obvious correlation between the minor variation in isotope data and other proxies of MS or biota.
