BIOSTRATIGRAPHY
Pleistocene through upper Maastrichtian sediments were recovered at Site 1262. Pliocene–Pleistocene assemblages show considerable reworking, and the Miocene through upper middle Eocene section yields calcareous microfossils strongly affected by carbonate dissolution as well as reworking. Lower Eocene and older sediments contain well-preserved calcareous assemblages. Shipboard examination of calcareous nannofossils and planktonic foraminifers permitted preliminary zonal and stage assignments (Fig. F22; Tables T5, T6). Benthic foraminifers place the sequence at lower abyssal depths during the late Paleocene through Pleistocene and at upper abyssal depths during the Cretaceous through early Paleocene. Biochronological ages plotted against mcd delineate overall sedimentation rates (Fig. F23) (see "Age Model and Mass Accumulation Rates").
Calcareous nannofossil assemblages were examined in the core catcher samples of all holes, and additional samples were inspected in numerous cores. Age and depth estimates of key biostratigraphic markers are shown in Table T5; a distribution chart showing the results from the core catcher samples is reported in Table T7.
Nannofossils are present in varying concentrations and states of preservation. In Cores 208-1262A-1H through 3H and Cores 208-1262B-1H through 5H, the assemblages have abundant nannofossils with good preservation. Sections 208-1262A-5H-CC and 6H-CC and 208-1262B-6H-CC, 7H-CC, and 9H-CC are barren. In sediments below 96 mcd (Cores 208-1262A-9H through 17H, 208-1262B-10H through 23H, and 208-1262C-1H through 14H), nannofossil assemblages are abundant, diverse, and generally well preserved, although nannofossils show varying degrees of dissolution and overgrowth.
Pleistocene–Oligocene (0–78 mcd)
The Pleistocene–Oligocene sequence (Core 208-1262A-1H through Section 7H-4 and Cores 208-1262B-1H through 9H) is characterized by slumps, turbidite layers, and thick intervals barren of calcareous nannofossils, making recognition of most nannofossil events of this time interval difficult. Moreover, assemblages are affected by reworking that increased anomalies in the biostratigraphic succession. On the basis of limited biostratigraphic data obtained from intermittent fossiliferous intervals, sediments from Cores 208-1262A-1H through 4H and Core 208-1262B-1H through Section 6H-2 were deposited during the Pleistocene–Pliocene. Lower Pliocene and upper Miocene nannofossil assemblages (Cores 208-1262A-5H and 208-1262B-6H) were extensively reworked and contain lower Miocene–Oligocene contaminants. Sediments in Cores 208-1262A-6H and 208-1262B-7H through 8H are barren of nannofossils, except for discrete layers that contain mixed assemblages of reworked Eocene forms.
Eocene/Oligocene Boundary Interval (75–81 mcd)
The carbonate-bearing sediments in the stratigraphic interval between 75 and 81 mcd (Cores 208-1262A-7H through 8H and 208-1262B-9H) were sampled to investigate the Eocene–Oligocene transition, including the uppermost occurrences of the last Paleogene representatives of rosette-shaped discoasters, Discoaster saipanensis and Discoaster barbadiensis. Nannofossil assemblages of Zones CP16 and CP15 (NP22, NP21, and NP20) straddle the Eocene–Oligocene transition. The boundary between Subzones CP16c and CP16b (NP22/NP21: top of Ericsonia formosa range) is distinct in both holes, whereas the uppermost occurrences (top [T]) of D. saipanensis and D. barbadiensis were recorded together and define the boundary between Subzone CP16a and Zone CP15 (NP21/NP20). The "cool-water taxon" Isthmolithus recurvus (Martini, 1971), whose range straddles the E/O boundary interval (from Zones CP16 through CP15; NP22 through NP19), is rare and has a discontinuous occurrence in Cores 208-1262A-8H and 7H and 208-1262B-9H. Major components of the lowermost Oligocene–uppermost Eocene assemblage are Reticulofenestra umbilicus, Reticulofenestra dictyoda, Dictyococcites spp., and Coccolithus eopelagicus. The absence of peaks in abundance of Ericsonia obruta, which characterize the E/O transition, suggests that this transition is incomplete at Site 1262.
Eocene (78–130 mcd)
The interval from 78 to 96 mcd (Sections 208-1262A-8H-1 through 9H-3 and 208-1262B-9H-6 through 10H-CC) is placed in the upper and middle Eocene. Assemblages characteristic of Zones CP15 to CP11 (NP19 to NP13) are recognized, but nannofossils are present in scattered samples only. From 84 to 94 mcd, assemblages belonging to Zones CP14, CP13, and CP12 (NP16, NP15, and NP14) are present along with common Sphenolithus spp., Chiasmolithus spp., Nannotetrina spp., and reworked lower Eocene species.
Nannofossils are abundant throughout the interval from 96 to 140 mcd. The uppermost part of this interval is in Zone CP11 (upper part of NP12) in the lower Eocene (Cores 208-1262A-9H, 208-1262B-11H, and 208-1262C-1H). Preservation varies from good to moderate, with overgrowths on discoasterids and dissolution of other forms. The main marker species for the lower Eocene zones are present (Table T7), including abundant Tribrachiatus orthostylus, Discoaster lodoensis, Sphenolithus radians, rare Discoaster multiradiatus, and common Discoaster diastypus. Most of the lower Eocene nannofossil zonal boundaries (from NP14/NP13 through NP12/NP11) have been recognized. In the Walvis Ridge region, Toweius crassus (marker of the CP11/CP10 boundary) seems to have a different stratigraphic range than indicated in the standard zonations; its range extends into Zone CP9 and was not used for the biozonation.
Paleocene/Eocene Boundary Interval (130–150 mcd)
Specimens belonging to the Rhomboaster-Tribrachiatus plexus are poorly preserved because of calcite recrystallization. They are present in Sample 208-1262A-13H-6, 25 cm, just above the benthic extinction event (BEE), but their distribution is not continuous in the overlying sediments, indicating that the lineage from Rhomboaster-Tribrachiatus to T. orthostylus is not fully represented at Site 1262. Overgrown specimens belonging to the Rhomboaster-Tribrachiatus plexus hindered identification of Tribrachiatus contortus and Tribrachiatus bramlettei. Thus, the boundaries between Subzones CP9b and CP9a (NP11/NP10) and between Subzone CP9a and Zone CP8 (NP10/NP9) could not be confidently recognized. The Subzone CP9b/CP9a (NP11/NP10) boundary is approximated by the appearance of S. radians. The genus Fasciculithus shows a distinct decrease in abundance in the uppermost part of its range. Fasciculiths are relatively common in the uppermost Paleocene but decrease in abundance, whereas Zygrhablithus bijugatus increases in abundance between Samples 208-1262A-13H-6, 10 cm, and 13H-6, 25 cm, just above the BEE. The uppermost occurrence of fasciculiths is between Samples 208-1262A-13H-4, 43 cm, and 13H-4, 60 cm. The reversal in relative abundances of the genera Fasciculithus and Zygrhablithus is prominent and easy to recognize and is observed just above the BEE. A similar "relative abundance reversal" between the genera Fasciculithus and Rhomboaster has been reported at equatorial Pacific Sites 1220 and 1221 (Lyle, Wilson, Janecek, et al., 2002) and may be correlative.
Paleocene (140–219 mcd)
Paleocene assemblages are generally diverse and moderately preserved because of slight dissolution of placoliths and some overgrowth on discoasterids. Important components of the assemblages are Toweius spp., Coccolithus pelagicus, Prinsius spp., Chiasmolithus spp., Cruciplacolithus spp., Heliolithus spp., and Ericsonia spp. Most of the key elements for the biostratigraphic classification of the interval from 153 to 219 mcd (Cores 208-1262A-14H through 17H, 208-1262B-16H through 22H, and 208-1262C-6H through 12H) were recorded with few exceptions (Table T7). The bottom of Heliolithus kleinpellii indicates the presence of the PBE between Samples 208-1262A-17H-2, 129 cm, and 17H-3, 34 cm, 208-1262B-18H-5, 40 cm, and 18H-5, 140 cm, and 208-1262C-9H-1, 150 cm, and Section 9H-CC. The marker of the Zonal CP7/CP6 boundary, Discoaster nobilis, was difficult to recognize because of overgrowth. The lowermost occurrence (bottom [B]) of Ellipsolithus macellus appears to be diachronous because its lowermost persistent occurrence is recorded in Zone CP5 (NP6), thereby preventing recognition of the Zonal CP3/CP2 (NP4/NP3) boundary.
Upper Cretaceous–Lower Paleocene Boundary Interval (211–232 mcd)
The K/P boundary interval is marked by a mass extinction of Cretaceous nannofossil taxa, together with a significant increase in abundance of Thoracosphaera spp., the B of Biantholithus sparsus (Sample 208-1262B-22H-4, 133 cm), and the co-occurrence of Markalius inversus and Cyclagelosphaera reinhardtii. These events are followed uphole by the appearance of new taxa such as Cruciplacolithus primus (64.8 Ma) and Cruciplacolithus tenuis s.s. (64.5 Ma). At Site 1262, the B of C. tenuis s.s. is between Samples 208-1262B-22H-2, 72 cm, and 22H-3, 33 cm, at an estimated depth of 213.79 mcd. The B of C. primus is between Samples 208-1262B-22H-3, 73 cm, and 22H-3, 130 cm, at an estimated depth of 214.45 mcd. An uppermost Maastrichtian nannofossil assemblage (Zone CC26) is present near the bottom part of the Site 1262 sedimentary succession. The B of Micula prinsii in Zone CC26 is recorded between Section 208-1262B-22H-CC and Sample 23H-1, 40 cm, at an estimated depth of 222.86 mcd.
Planktonic foraminifers were examined in all core catcher samples from Holes 1262A and 1262B, as well as at a higher resolution across the P/E and K/P boundary intervals (see Tables T6, T8).
Pleistocene–Oligocene (0–78 mcd)
Pleistocene assemblages preserved in Cores 208-1262A-1H and 208-1262B-1H through 2H (0.62–12.31 mcd) consist of a mixture of well-preserved subtropical and temperate planktonic foraminifers. The uppermost part of Core 208-1262B-1H may contain Holocene sediments, but this was not possible to confirm. Common species are Globorotalia crassaformis, Globorotalia truncatulinoides, Globorotalia tumida, Globorotalia ungulata, Globoconella inflata, Globigerinoides ruber, Globigerinoides sacculifer, Globigerina falconensis, Globigerina (Zeaglobigerina) rubescens, Globigerinella siphonifera, Hirsutella scitula, and Neogloboquadrina pachyderma (dextral). The T of Globorotalia tosaensis (base of Subzone Pt1b; 0.65 Ma) is in Sample 208-1262B-1H-4, 62–64 cm (~4.65 mcd). The base of Subzone Pt1a (~2.02 Ma) is used to approximate the Pliocene/Pleistocene boundary and is placed at ~19.20 mcd (Sample 208-1262A-1H-2, 62–64 cm, and Section 208-1262B-2H-CC). The lower portion of the Pleistocene (Section 208-1262A-1H-6) has been extensively reworked (Table T8).
Many of the tropical/subtropical age-diagnostic taxa used to subdivide the upper Miocene–Pliocene interval are missing from the section because of temperate environmental conditions. Preservation is variable, ranging from poor to good (Table T8). A reliable biozonation could not be constructed because of pervasive reworking, but specimens transitional between Globoconella conomiozea and Globoconella puncticulata (i.e., Globoconella sphericomiozea) are present in Section 208-1262B-5H-CC, providing a loose approximation for the Miocene/Pliocene boundary at ~43 mcd. The depth of the G. crassaformis lowermost occurrence (48.66 mcd) plots as an outlier along the age-depth curve, suggesting that this datum is diachronous (Fig. F23).
The interval from Sections 208-1262A-4H-CC through 6H-CC (40–68 mcd) contains a condensed upper Miocene through upper Oligocene section. Assemblages from this stratigraphic interval are very poorly preserved because of intense carbonate dissolution and fragmentation; only thick-shelled taxa resistant to dissolution were found intact (Table T8). The presence of robust Sphaeroidinellopsis disjuncta and Globigerina druryi/Globigerina nepenthes indicates a middle Miocene age for Section 208-1262A-5H-CC (56.83 mcd). Some carbonate-rich turbidite layers in Cores 208-1262A-5H and 6H contain well-preserved middle Eocene taxa (e.g., Acarinina bullbrooki). The stratigraphic position of the Oligocene/Miocene boundary could not be discerned, and the upper Oligocene may be missing (Fig. F22).
Section 208-1262A-7H-CC (77.92 mcd) is loosely constrained to the lower Oligocene based on the co-occurrence of Subbotina angiporoides, Catapsydrax unicavus, Globorotaloides suteri, and Chiloguembelina cubensis. Test preservation in these assemblages remains relatively poor, with only thick-shelled dissolution-resistant taxa present.
Eocene/Oligocene Boundary Interval (78–88 mcd)
The E/O boundary transition was not recognized at the core catcher sampling resolution. Intense carbonate dissolution and a dearth of tropical marker taxa hindered precise age determination for Section 208-1262A-8H-CC (87.68 mcd), but the co-occurrence of Globigerinatheka spp. and Acarinina spp. and the absence of Morozovella aragonensis restricts the age of this sample to the middle Eocene (foraminiferal Zones P12–P14). Thus, the entire upper Eocene is highly condensed or contains unconformities confined to an interval that is only ~10 m thick.
Lower Eocene (~90–140 mcd)
Preservation improves downhole as lower Eocene planktonic foraminiferal assemblages are better preserved than those in the overlying units. The presence of M. aragonensis and the absence of such common middle Eocene taxa as A. bullbrooki and Acarinina spinuloinflata constrain Section 208-1262A-9H-CC (100.84 mcd) to Zones P7 and P8. The scarcity of Morozovella formosa—a critical marker species—prevents a more precise biostratigraphic assignment for Section 208-1262A-9H-CC, although Zone P7 (52.30–50.80 Ma) is more consistent with the co-occurring calcareous nannoflora (Fig. F22).
Planktonic foraminifers in Sections 208-1262A-10H-CC through 12H-CC (111–142 mcd) are moderately to well preserved and are assigned to Zone P6; subdivision of this zone was precluded by the absence of M. formosa. The diverse assemblages are largely dominated by species belonging to the genus Acarinina with subordinate numbers of morozovellids, subbotinids, globanomalinids, chiloguembelinids, and igorinids. Relatively large specimens of Chiloguembelina sp. are present in Section 208-1262B-12H-CC (115.69 mcd).
Paleocene/Eocene Boundary Interval (139.5–142 mcd)
Samples were taken at ~10-cm spacings across the P/E boundary interval in Core 208-1262A-13H. In general, the Morozovella velascoensis group is poorly represented in the P/E boundary interval at Site 1262 (Table T8). The T of M. velascoensis is provisionally placed in Sample 208-1262A-13H-5, 138–139 cm (139.49 mcd). The stratigraphic range of M. velascoensis typically extends well into the lowermost Eocene, but at Site 1262, its range is abbreviated primarily because of suboptimal ecological conditions. Samples 208-1262A-13H-5, 138–139 cm (139.49 mcd), through 13H-6, 19–20 cm (139.80 mcd), contain abundant moderately preserved planktonic foraminifers. Primary constituents of these assemblages are Morozovella subbotinae, Morozovella aequa, Acarinina soldadoensis, Acarinina coalingensis, chiloguembelinids, Globanomalina australiformis, and subbotinids.
The overall abundance and preservation of planktonic foraminifers declines downhole to the P/E boundary. Samples 208-1262A-13H-6, 30–31 cm (139.91 mcd), and 43–44 cm (140.04 mcd), are from a clay-rich layer that contains few planktonic foraminifers. These depauperate assemblages are moderately preserved and consist of diminutive acarininids, subbotinids, and morozovellids. Sample 208-1262A-13H-6, 57 cm (140.18 mcd), also from the clay layer, yields an assemblage composed of relatively small acarininids, morozovellids, chiloguembelinids, and rare G. australiformis. Unlike other low- to mid-latitude P/E boundary intervals, the Site 1262 record contains very few M. velascoensis and/or related "excursion" taxa (e.g., Kelly et al., 1996); only one specimen of Morozovella allisonensis was found.
Uppermost Paleocene Sample 208-1262A-13H-6, 95 cm (140.56 mcd), and Section 13H-CC (142.08 mcd) from below the clay layer contain moderately preserved foraminifers that exhibit a relatively high degree of fragmentation. The presence of rare, diminutive forms of M. velascoensis places these two samples in Zone P5.
Upper Paleocene (140–193.5 mcd)
Preservation among upper Paleocene assemblages is variable but generally moderate. The top of Zone P4, as delimited by the T of Globanomalina pseudomenardii, falls between Sections 208-1262B-15H-CC and 16H-CC at an estimated depth of ~151 mcd (Fig. F22). The bottom of Zone P4, as delimited by the B of G. pseudomenardii, falls between Sections 208-1262B-18H-CC and 19H-CC at an estimated depth of ~183 mcd. The B of M. velascoensis occurs between Sections 208-1262B-18H-CC and 19H-CC at an estimated depth of ~183 mcd. The boundary between the upper and lower Paleocene (Danian) is correlative to the base of Subzone P3a and is denoted by the B of Morozovella angulata, which occurs between Sections 208-1262B-19H-CC and 20H-CC at an estimated depth of ~193.5 mcd.
Upper Cretaceous–Lower Paleocene Boundary Interval (193.5–217 mcd)
Core catcher samples bracketing the K/P boundary were supplemented by sampling from Core 208-1262B-22H. Assemblages are generally well preserved, although the Maastrichtian faunas exhibit varying degrees of etching and dissolution. The base of Zone P2 is placed between Sections 208-1262B-20H-CC and 21H-CC and is thereby constrained to ~204.5 mcd. Zone P2 is underlain conformably by Subzone P1c, which is estimated to extend from ~204.64 mcd to ~214.62 mcd (Fig. F22). Planktonic foraminifers in this expanded P1c–P2 interval are well preserved.
"Dwarfed" assemblages from Subzones P1a and P1b are generally well preserved and consist primarily of diminutive forms belonging to the following taxa: Praemurica taurica, Parasubbotina pseudobulloides, Globanomalina archeocompressa, Globanomalina planocompressa, Globoconusa daubjergensis, Eoglobigerina eobulloides, and Guembelitria cretacea. The base of Subzone P1b, as delimited by the B of Subbotina triloculinoides, is tentatively placed at 216 mcd (Fig. F22).
The zonal boundary between Subzone P1a and Zone P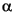 is located between Samples 208-1262B-22H-4, 86 cm, and 22H-4, 115 cm, at an estimated depth of ~216 mcd. Large reworked specimens of Maastrichtian foraminifers are present in Zone P, whereas Zone P0 was not recognized in the Hole 1262B K/P boundary section. The presence of Abathomphalus mayaroensis and Contusotruncana contusa in Sample 208-1262B-22H-4, 144 cm (216.58 mcd), signifies the Maastrichtian. Thus, the K/P boundary (Fig. F22) is situated between Samples 208-1262B-22H-4, 134 cm (216.48 mcd), and 22H-4, 144 cm (216.58 mcd).
is located between Samples 208-1262B-22H-4, 86 cm, and 22H-4, 115 cm, at an estimated depth of ~216 mcd. Large reworked specimens of Maastrichtian foraminifers are present in Zone P, whereas Zone P0 was not recognized in the Hole 1262B K/P boundary section. The presence of Abathomphalus mayaroensis and Contusotruncana contusa in Sample 208-1262B-22H-4, 144 cm (216.58 mcd), signifies the Maastrichtian. Thus, the K/P boundary (Fig. F22) is situated between Samples 208-1262B-22H-4, 134 cm (216.48 mcd), and 22H-4, 144 cm (216.58 mcd).
Core catcher samples from Hole 1262A were semiquantitatively investigated for benthic foraminifers, as were core catchers from Hole 1262B for intervals not recovered in Hole 1262A and additional samples around critical intervals (Table T9).
In most samples, benthic foraminifers are rare and strongly outnumbered by planktonic foraminifers, with the exception of samples in which strong dissolution has affected the foraminiferal assemblages. In these samples, benthic foraminifers are common, but planktonic foraminifers are mostly absent, and there is a residue of fish debris (Sections 208-1262A-7H-CC, 8H-CC, 16H-CC, and 17H-CC and interval 208-1262C-9H-1, 145–157 cm). These samples contain evidence of downslope transport. Sections 208-1262A-6H-CC and 208-1262B-9H-CC and Sample 208-1262A-13H-6, 43–44 cm, are barren.
Benthic foraminiferal assemblages from Site 1262 indicate deposition at lower abyssal depths (>3000 m) for Sections 208-1262B-1H-CC through 17H-CC (0–166 mcd). Assemblages in samples from 166 through 188 mcd (Sections 208-1262B-17H-CC through 19H-CC) indicate deposition at transitional upper to lower abyssal depths (e.g., Alegret and Thomas, 2001), and assemblages below 188 mcd indicate lowermost upper abyssal depths (~3000 m).
The mudline sample and core catcher samples down through interval 208-1262A-4H-CC (46 mcd) contain faunas similar to those presently living in the Walvis Ridge area (Mackensen et al., 1995; Schmiedl et al., 1997), with rare Epistominella exigua and Alabaminella weddellensis (phytodetritus-consuming species). Assemblages are dominated by Nuttallides umbonifera, an indicator of the presence of Antarctic Bottom Water, which has been argued to be present to the north of Walvis Ridge below depths of ~4200 m (Schmiedl et al., 1997). Cibicidoides wuellerstorfi is rare between 0 and 36 mcd. Other common species include Pullenia spp., Oridorsalis umbonatus, Stainforthia complanata, and Globocassidulina subglobosa.
Assemblages in Sections 208-1262A-5H-CC (56.83 mcd) through 208-1262B-9H-CC (83.94 mcd) are difficult to interpret because of strong dissolution, leaving only solution-resistant forms and signs of downslope transport of large, heavy-shelled taxa.
Section 208-1262A-9H-CC (100.84 mcd) through Sample 13H-6, 30–31 cm (139.91 mcd), generally contain rare benthic foraminifers because of very high planktonic to benthic ratios. The faunas are similar to those described at other abyssal sites in the South Atlantic Ocean (Clark and Wright, 1984; Müller-Merz and Oberhänsli, 1991; Thomas and Shackleton, 1996), with common to abundant small smooth-walled species belonging to the genera Abyssamina, Quadrimorphina, and Clinapertina and common to abundant Nuttallides truempyi. Other taxa commonly present include Tappanina selmensis, Siphogenerinoides brevispinosa, and Aragonia aragonensis (none of the three are more abundant than a few percent), various species of Gyroidinoides, laevidentalinid and pleurostomellid taxa, Anomalinoides spissiformis, Nonion havense, and O. umbonatus. The species richness fluctuates, with the abundance of various abyssaminid taxa inversely correlated to species richness. The lowest species richness and a high relative abundance of Abyssamina poagi occur in the lower part of the interval and in Sample 208-1262C-2H-6, 35–36 cm (117.14 mcd), in a reddish clay layer.
The major extinction of benthic foraminifers at the end of the Paleocene (BEE) occurs between Sample 208-1262A-13H-6, 57–58 cm (140.18 mcd), and barren Sample 208-1262A-13H-6, 43–44 cm (140.04 mcd). Faunas in the lowermost nonbarren samples (i.e., immediately after the extinction) have very low species richness and are dominated by small thin-walled specimens of N. truempyi and various species of Abyssamina, Clinapertina, and Quadrimorphina, as at DSDP Site 527 (Thomas and Shackleton, 1996). The uppermost sample below the extinction has a very diverse fauna with a typical upper Paleocene assemblage, but all specimens are diminutive.
Samples between 208-1262A-13H-6, 57–58 cm, through 15H-CC (140–162 mcd) contain a typical preextinction Paleocene highly diverse abyssal assemblage, with common Stensioeina beccariiformis, Cibicidoides hyphalus, Pullenia coryelli, Bulimina thanetensis, Aragonia velascoensis, large agglutinative taxa such as Clavulinoides amorpha, Clavulinoides trilatera, Marssonella oxycona, as well as large smooth-walled Gyroidinoides species such as Gyroidinoides beisseli and Gyroidinoides globosus, all of which had their last appearance during the extinction. In this interval, S. brevispinosa and Rectobulimina carpentierae are present in most samples and their relative abundances are highly variable. Lower bathyal indicator species are absent.
The assemblages change gradually in species composition in the interval between Sections 208-1262B-17H-CC and 19H-CC (166–188 mcd), with Abyssamina and Clinapertina species as well as B. thanetensis becoming less common downhole and Nuttallinella florealis and Nuttallinella sp. becoming more common. At the bottom of this interval, Spiroplectammina dentata has its local uppermost occurrence, as does Nuttallinella coronula and Tritaxia havanensis. Benthic foraminiferal assemblages changed at about this time at ODP Sites 689 and 690 at Maud Rise (Thomas, 1990), but at Site 1262, the faunal changes might be the result of a change in paleodepth from upper abyssal in the lower part of the section to lower abyssal in its upper parts.
The benthic foraminiferal assemblages below 188 mcd closely resemble assemblages in the Velasco Formation in Mexico, where many of the species present at Site 1262 were first described (see review in Alegret and Thomas, 2001). Assemblages across the K/P boundary (Samples 208-1262B-22H-4, 134–135 cm, and 22H-CC; 216.50–221.75 mcd) document that benthic foraminifers at Site 1262 do not suffer significant extinction, as is observed at many other sites. The uppermost occurrences of Angulogerina szajnochae and Praebulimina reussi occur close to the K/P boundary in the preliminary data, but it is not well documented whether these species disappeared globally at that time.
