BIOSTRATIGRAPHY
Abyssal depths below the CCD characterize the environment of the Paleogene and Cretaceous section cored at Site 1276. Consequently, carbonate microfossil assemblages, particularly the foraminifers, were significantly impacted by dissolution. Calcareous nannofossils, owing to their transport to the seafloor in zooplankton fecal pellets (Honjo, 1975, 1976; Pilskaln and Honjo, 1987; Steinmetz, 1994), fared much better against the ravages of dissolution than the calcareous foraminifers. The nannofossils show variable abundance and preservation through the section but provide excellent biostratigraphic control in most sampled intervals. Palynomorphs (predominantly dinocysts and sporomorphs) are very well preserved in carbon-rich, dark-colored sediments, and they provide a critical component of the biostratigraphic age control for the site.
Planktonic foraminifers are generally rare or absent in shale and mudrock facies at this site, but they are typically present in turbiditic units and provide complementary biostratigraphic control through much of the Paleogene and Cretaceous. Most foraminiferal samples (>63-µm residues) contain fairly diverse and generally well preserved assemblages of in situ abyssal (>2000 m) agglutinated benthic foraminifers. A summary of the microfossil biostratigraphy is compared to the lithology and core recovery in Figure F140, and major micropaleontological features of the shipboard core catcher samples and thin sections are summarized in Table T7.
Where present, planktonic and calcareous benthic foraminifers are often very small (<125 µm). They likely are size sorted by processes associated with downslope transport in turbidity currents, a supposition that is based on the similar size of co-occurring quartz grains. Calcareous benthic foraminifers indicate that the source areas for the turbidites cored at Site 1276 have varied through the section from neritic (<200 m) to bathyal (200-2000 m) depths along the adjacent continental margin. Reworking of Upper Cretaceous material is prevalent in the Paleogene, but redeposition processes were largely penecontemporaneous during the Cretaceous, based on a uniform age progression of the samples and minimal reworking of older material in calcareous nannofossil, palynomorph, and foraminifer samples.
An unconformity separates lithologic Units 1 and 2 in Core 210-1276A-8R, based on palynomorph and calcareous nannoplankton evidence, and two condensed intervals are identified in the Upper Cretaceous (lower to middle Maastrichtian in Core 22R, and uppermost Turonian-Coniacian to upper Campanian in Cores 25R through 28R). Core 210-1276A-14R crosses the PETM but did not recover the dissolution interval that characterizes this event. The K/T boundary interval may be biostratigraphically complete in Section 210-1276A-21R-4 based on calcareous nannofossil and planktonic foraminiferal evidence. It is likely that Cenomanian-Turonian OAE 2 ("Bonarelli" event) is represented by the laminated black shales in Sections 210-1276A-31R-2 and 31R-3, as constrained by the shipboard biostratigraphy. Basal Albian OAE 1b ("Paquier" event) was not conclusively defined by routine shipboard biostratigraphy, but it likely occurs above the lower sill in Core 210-1276A-94R, based on supporting geochemical data. Other apparent OAEs are also outlined in "Oceanic Anoxic
Events" in "Geochemistry."
Calcareous nannofossils, dinocysts, and planktonic foraminiferal assemblages indicate that Site 1276 was influenced by transitional surface water masses for much of its Paleogene and Cretaceous history. This is indicated by the presence of select high-latitude taxa and by the absence or paucity of some key low-latitude Tethyan taxa.
Calcareous nannofossils recovered at Site 1276 exhibit a wide range of preservation and abundance. Abundance and preservation are usually best in the tops of the sandy turbidites found throughout the drilled section. The hemipelagic sediments generally either contain very poorly preserved and impoverished assemblages or they are completely barren, indicating deposition well below the CCD.
Nannofossil distribution in Hole 1276A was found to be discontinuous because of the dissolution of carbonates. Nonetheless, the distribution allowed a fairly complete biostratigraphic analysis. However, the dominance of redeposited sediments and the common presence of reworked taxa make the use of last occurrence (LO) datums problematic.
Paleogene
Lithologic Units 1-3 consist of hemipelagic mudrocks interrupted by turbidites and debris flows. The redeposited sediments are the only lithologies suitable for calcareous nannofossil analysis because the mudstones are completely devoid of calcium carbonate.
The interval between Samples 210-1276A-1W-CC and 2R-CC contains rare to abundant, moderately well preserved nannofossils with common reworked taxa from older Paleogene and Cretaceous strata. Samples 210-1276A-1W-4, 37 cm, and 1W-CC belong to uppermost Eocene-lower Oligocene Zone CP16, based on the presence of Reticulofenestra umbilicus, R. bisecta, R. oamaroensis, Isthmolithus recurvus, and Chiasmolithus altus and the absence of "rosette" discoasters (D. barbadiensis and D. saipanensis) (Table T8). Also present in these samples are moderately well preserved specimens of Chiasmolithus expansus, Pemma basquensis, P. papillatum, Coccolithus formosus, and Blackites spinosus. Specimens of Cribrocentrum reticulatum and very rare Discoaster saipanensis are considered to be reworked.
Sample 210-1276A-2R-1, 144 cm, is assigned to upper Eocene Zone CP15 based on the presence of Reticulofenestra oamaroensis. Sample 210-1276A-2R-CC is assigned to undifferentiated Subzone CP14b-Zone CP15 based on the presence of Reticulofenestra bisecta, R. umbilicus, and C. grandis. Many of the larger clasts in the debris flows in this core (Samples 210-1276A-2R-2, 83 cm, and 2R-3, 36 cm) contain well-preserved and diverse assemblages of older middle Eocene taxa (Zone CP13). Samples 210-1276A-3R-2, 51 cm, and 3R-CC are assigned to middle Eocene Subzone CP14a based on the presence of Chiasmolithus solitus occurring with R. umbilicus. Samples 210-1276A-4R-CC through 7R-CC are barren of nannofossils.
Samples 210-1276A-8R-CC through 11R-CC yield fairly abundant and moderately to poorly preserved assemblages. There is a marked difference in the preservation of nannofossils between lithologic Units 1 and 2, with preservation being much better in the former. The presence of Rhabdosphaera inflata together with questionable Nannotetrina fulgens in Sample 210-1276A-8R-CC indicates lower middle Eocene Subzone CP12b or, perhaps, Subzone CP13a. Also present in this sample are Discoaster lodoensis, D. sublodoensis, Ellipsolithus macellus, and E. lajollaensis. The absence of R. inflata in Sample 210-1276A-9R-CC indicates Subzone CP12a.
Samples 210-1276A-10R-CC and 11R-CC are assigned to the undifferentiated lower Eocene Zones CP10-CP11 based on the presence of D. lodoensis in the absence of D. sublodoensis. Zones CP10 and CP11 could not be differentiated because Toweius crassus was not observed in these sediments. However, Sample 210-1276A-12R-4, 55 cm, may be constrained to lower Eocene Zone CP10 based on a few occurrences of Tribrachiatus orthostylus. Sample 210-1276A-12R-CC is barren of nannofossils, and Sample 13R-CC contains only rare specimens of Discoaster multiradiatus.
The Paleocene/Eocene Boundary
The Paleocene/Eocene (P/E) boundary is characterized by an abrupt worldwide warming event referred to as the PETM. In the oceanic sedimentary record, the PETM interval typically corresponds to a sharp lithologic change uphole from light calcareous marl to a dark, clayey, calcite-free dissolution interval a few centimeters to decimeters thick. This interval is also distinguished by a sharp negative  13C excursion and a benthic foraminiferal extinction event (Zachos et al., 1993; Thomas and Shackleton, 1996). Biostratigraphically, the PETM lies at the top of calcareous nannofossil Zone CP8. At the Paleocene-Eocene transition, the calcareous nannoplankton community shows a great turnover characterized by peculiar biotic changes: the first members of the Rhomboaster-Tribrachiatus lineage (Rhomboaster spp. and R. cuspis) are present just above the dissolution interval and are followed by a sharp decrease in fasciculithid abundance and an increase in Zygrhablithus bijugatus abundance (Bralower et al., 1995). During this time interval until the recovery, the calcareous nannofossil assemblages are dominated by the genus Discoaster.
13C excursion and a benthic foraminiferal extinction event (Zachos et al., 1993; Thomas and Shackleton, 1996). Biostratigraphically, the PETM lies at the top of calcareous nannofossil Zone CP8. At the Paleocene-Eocene transition, the calcareous nannoplankton community shows a great turnover characterized by peculiar biotic changes: the first members of the Rhomboaster-Tribrachiatus lineage (Rhomboaster spp. and R. cuspis) are present just above the dissolution interval and are followed by a sharp decrease in fasciculithid abundance and an increase in Zygrhablithus bijugatus abundance (Bralower et al., 1995). During this time interval until the recovery, the calcareous nannofossil assemblages are dominated by the genus Discoaster.
The search for the P/E boundary at Site 1276 was difficult for two important reasons: (1) the presence of numerous clayey, calcite-free intervals deposited below the CCD and (2) the occurrence of turbidites and debris flows, which blur the typically distinct lithologic and paleontological record, specifically, the absence of a single dissolution interval and the reworking of key taxa. To better constrain the placement of the PETM and the P/E boundary at Site 1276, a preliminary analysis was conducted on samples spanning the interval from Sections 210-1276A-13R-1 through 15R-3. The samples from Core 210-1276A-13R contain low-abundance and low-diversity assemblages dominated by Discoaster spp. and Rhomboaster spp., and they lack fasciculithids. These assemblages are affected by dissolution, which hinders the recognition of key species. The highest Tribrachiatus orthostylus specimen occurs in Sample 210-1276A-13R-1, 35 cm, indicating lower Eocene Subzone CP9b. The first occurrence (FO) of T. contortus occurs in Sample 210-1276A-13R-1, 96 cm; this event lies near the base of Subzone CP9a, which is defined by the FOs of Discoaster diastypus and Tribrachiatus bramlettei in Sample 14R-1, 24-25 cm. The LO of Fasciculithus tympaniformis occurs in Sample 210-1276A-14R-1, 104-105 cm; few to rare fasciculiths are recorded from this sample downhole to Sample 14R-CC, which contains a diverse assemblage of nannofossils, including common fasciculithids. Almost all samples from Core 210-1276A-14R are characterized by very poorly preserved, low-diversity, and low-abundance assemblages. The most common taxa in these poor assemblages are Discoaster spp. and Rhomboaster spp. These preliminary results suggest that the recovered sediments of Sections 210-1276A-14R-1 to 14R-4 contain only the characteristic post-PETM-excursion calcareous nannofossil assemblages and succession of biotic events. Therefore, the PETM dissolution interval likely occurred in the unrecovered interval between Samples 210-1276A-14R-4, 105-106 cm, and 14R-CC.
Sample 210-1276A-15R-2, 25-27 cm, is assigned to upper Paleocene Zone CP8 based on the presence of Discoaster multiradiatus and Chiasmolithus bidens and the absence of typical Eocene taxa. Sample 210-1276A-15R-CC is barren of nannofossils, but Samples 16R-CC and 17R-CC contain low-diversity and poorly preserved calcareous nannofossil assemblages with common reworked Cretaceous species. The marker for mid-Paleocene Zone CP3, Ellipsolithus macellus, was not found but is known to be dissolution prone. These samples are nonetheless assigned to Zone CP3 because of the presence of Fasciculithus ulii in Sample 210-1276A-16R-CC and F. magnus in Sample 17R-CC. Chiasmolithus danicus, which fixes the base of lower Paleocene Zone CP2, is common in both of these samples. Sample 210-1276A-18R-CC is assigned to an undifferentiated Zone CP2-CP3 based on the absence of any Fasciculithus species and the presence of C. danicus. The missing Zones CP4-CP7 may be differentiated with more detailed sampling of Cores 210-1276A-15R and 16R.
Samples 210-1276A-19R-CC and 20R-CC are barren; however Sample 20R-5, 104-105 cm, contains Cruciplacolithus primus, Biantholithus sparsus, common Markalius inversus, and Neocrepidolithus neocrassus, together with numerous fragments of Thoracosphaera spp. This assemblage is typical of basal Paleocene Subzone CP1a.
The Cretaceous/Tertiary Boundary
Determining the position of the K/T boundary transition in Site 1276 was difficult because of extensive reworking and the presence of several intervals of carbonate-free sediments. In several low-latitude localities that contain a biostratigraphically continuous transition between the uppermost Maastrichtian and the lowermost Danian, the K/T boundary is marked by peculiar biotic changes in the calcareous nannoplankton community (Gardin and Monechi, 1998, and references therein). These changes include the following: the lowest Danian sediments are characterized by a dramatic increase in the abundance of the calcareous dinocyst Thoracosphaera spp. ("Thoracosphaera bloom"), occurring together with abundant Cretaceous species that are considered to be reworked. This first bloom of Thoracosphaera spp. is followed by an increase in abundance of survivor species such as Markalius inversus, Cyclagelosphaera reinhardtii, and Braarudosphaera bigelowii and by blooms of dwarf incoming Danian species such as Neobiscutum romeinii and N. parvulum ("dwarf Biscutum bloom;" Gardin and Monechi, 1998). The first more complex Danian species, Cruciplacolithus primus, appears during this time and rapidly increases in size and abundance.
All these biotic signals help to constrain the K/T boundary transition at Site 1276 to Section 210-1276A-21R-4. Samples 210-1276A-21R-4, 53-54 cm, and 21R-4, 77 cm, contain abundant and diverse uppermost Maastrichtian assemblages belonging to the Micula prinsii Subzone UC20d. No thoracosphaerid fragments or unusually high abundances of survivor species are found in these samples, and as such, they are considered to be in situ. Sample 210-1276A-21R-4, 56-57 cm, is from a dark brownish green clay layer and yields only very rare, dissolution-resistant Cretaceous species such as Micula decussata and Watznaueria barnesae. Sample 210-1276A-21R-4, 49 cm, from the carbonate turbidite overlying this clay layer, also contains abundant and diverse Maastrichtian assemblages with no apparent Danian biotic components. Sample 210-1276A-21R-4, 41 cm, still contains abundant Maastrichtian species but has Danian thoracosphaerid fragments and is interpreted to be in situ. Samples 210-1276A-21R-2, 26 cm, and 21R-3, 52 cm, are also characterized by abundant Cretaceous species, with frequent Thoracosphaera spp. and very abundant Neobiscutum parvulum and N. romeinii. This succession of assemblages is comparable to that observed elsewhere at the K/T transition, even though the abundance of Cretaceous species above the transition at Site 1276 is remarkably high due to redeposition. Based on the calcareous nannofossil assemblages, the K/T boundary can be biostratigraphically placed in Section 210-1276A-21R-4 between 41 and 49 cm. However, it is possible that the Maastrichtian assemblage found in Sample 210-1276A-21R-4, 49 cm, consists of entirely reworked fossils in a barren lower Danian clay layer.
Upper Cretaceous
Sample 210-1276A-21R-CC yields a poorly preserved assemblage that is characterized here as upper Maastrichtian Zone UC20 based on the presence of M. prinsii (Table T9). Sample 210-1276A-22R-1, 2 cm, is better preserved and indicates the presence of upper Maastrichtian Subzones UC20a-UC20c based on the lack of M. prinsii and the presence of common Lithraphidites quadratus, Cribrocorona gallica, Arkhangelskiella maastrichtiana, and Nephrolithus frequens. The assemblage in Sample 210-1276A-22R-2, 50 cm, is characterized by the absence of L. quadratus and Tranolithus orionatus, which could indicate undifferentiated mid-Maastrichtian Zones UC19-UC18. Sporadic occurrences of Reinhardtites levis and Uniplanarius trifidus in this sample are considered to be reworked. Samples 210-1276A-22R-CC through 25R-5, 48-49 cm, contain common R. levis, U. trifidus, and Broinsonia parca constricta, which indicate mid- to upper Campanian Zones UC15-UC16. These results imply that lower Maastrichtian Zone UC17 is missing or difficult to distinguish because of reworking. It is likely that much of the Maastrichtian at this site is condensed in Core 210-1276A-22R.
The upper Campanian and upper Maastrichtian assemblages are characterized by typical low-latitude Tethyan taxa such as U. trifidus and L. quadratus, mixed with cooler-water high-latitude species such as Cribrosphaerella daniae and Psyktosphaera firthii. Nephrolithus frequens, the marker taxon of upper Maastrichtian Zone CC26 (Sissingh, 1977), is common, whereas the tropical species Micula murus (a marker for this zone at low latitudes) is very rare at Site 1276. These data suggest that Site 1276 occupied an intermediate region between the Boreal and Tethyan provinces and that mixed assemblages were common during this time period.
Sections between 210-1276A-25R-6 and 28R-5 are barren of calcareous microfossils. Many samples were taken from different lithologies in this thick interval in an attempt to constrain the age, but none yielded nannofossils. It is assumed that the interval contains all of the zones between mid-Campanian Zone UC15 and upper Coniacian Zone UC10. Samples 210-1276A-28R-5, 76-77 cm, and 28R-CC contain a rather depauperate assemblage assigned to uppermost Turonian to lower Coniacian Zone UC9, based on the absence of Broinsonia spp., Arkhangelskiella cymbiformis, and Micula decussata and on the presence of Eiffellithus eximius, Quadrum gartneri, Lithastrinus septenarius, Kamptnerius magnificus, and Marthasterites furcatus. This age assignment indicates that most of the Coniacian through the lower Campanian is either missing or is condensed in the barren red sandstones to mudstones of Cores 210-1276A-25R through 28R.
Sample 210-1276A-29R-CC is middle to upper Turonian Zone UC8 based on the presence of Eiffellithus eximius, and Sample 30R-CC is lower Turonian Zone UC7 based on the presence of Eprolithus moratus, E. octopetalus, and Quadrum gartneri. Samples 210-1276A-31R-CC and 32R-CC are barren, but Samples 31R-1, 25-26 cm, and 31R-4, 120-121 cm, yield moderately well preserved and abundant assemblages that help constrain the placement of the Cenomanian/Turonian boundary. The FO of E. octopetalus is recorded in Sample 210-1276A-31R-1, 25-26 cm, indicating an early Turonian age, whereas Samples 31R-4, 120-121 cm, and 33R-CC are assigned to undifferentiated Cenomanian Zone UC3-UC5 based on the presence of Quadrum intermedium and the absence of E. octopetalus. Unfortunately, the common presence of reworked taxa prevents the reliable recognition of important last occurrences that characterize the upper Cenomanian to lower Turonian transition, including Lithraphidites acutus, Corollithion kennedyi, and Helenea chiastia. Samples 210-1276A-30R-CC through 34R-CC are also characterized by fairly high values of TOC. These results, in conjunction with the planktonic foraminiferal biostratigraphic data, indicate the likely presence of the OAE 2 "Bonarelli event" in Sections 210-1276A-31R-2 and 31R-3.
Samples 210-1276A-34R-CC and 35R-CC are placed into upper Cenomanian Zone UC3 based on the presence of frequent Lithraphidites acutus and the absence of Cylindralithus biarcus. This assemblage is also characterized by common Axopodorhabdus albianus, Eiffellithus turriseiffelii, Helicolithus compactus, Rhagodiscus achlyostaurion, R. asper, and Corollithion kennedyi.
Sample 210-1276A-36R-CC is assigned to lower Cenomanian Zone UC1 based on the presence of C. kennedyi in the absence of L. acutus and Gartnerago segmentatum. Samples 210-1276A-37R-CC through 44R-CC are placed into upper Albian-basal Cenomanian Subzone NC10a based on the absence of C. kennedyi and the continued presence of E. turriseiffelii. Samples 210-1276A-39R-CC, 41R-CC, and 42R-CC are barren of nannofossils.
Lower Cretaceous
The Albian/Cenomanian boundary is straddled by Subzone NC10a, which is discussed in the section above. Upper Albian Subzone NC9b persists from Samples 210-1276A-45R-CC through 51R-CC based on the presence of Eiffellithus monechiae and the absence of E. turriseiffelii. In this interval, Sample 210-1276A-50R-CC (black mudstone) is barren of nannofossils. Sample 210-1276A-52R-CC is also nearly barren but may belong to this zone because of the rare occurrence of Eiffellithus cf. E. monechiae in Sample 54R-CC. However, these specimens may be ancestral forms.
Beginning with Sample 210-1276A-53R-CC and persisting through 75R-CC, the very rare occurrence of Axopodorhabdus albianus in the absence of E. monechiae indicates that this interval belongs to middle Albian Subzone NC9a. Samples 210-1276A-56R-CC through 58R-CC and 61R-CC, 66R-CC, 68R-CC, and 70R-CC through 72R-CC are barren of calcareous nannofossils. Throughout this part of the section there are occasional occurrences of Hayesites albiensis, Eprolithus floralis, Cylindralithus nudus, Braarudosphaera stenoretha, Lithraphidites carniolensis, and nannoconid species such as Nannoconus truitti, N. wassallii, and N. steinmannii.
Sample 210-1276A-76R-CC is assigned to middle Albian Subzone NC8c based on the presence of Tranolithus orionatus and H. albiensis, together with Prediscosphaera columnata. In the interval from Sample 210-1276A-77R-CC to 90R-CC, a number of core catcher samples are either barren of calcareous nannofossils (Table T9) or the absence of secondary marker taxa prohibits consistent subdivision of Zone NC8. Uppermost Aptian-lower Albian Subzone NC8a is assigned to Samples 210-1276A-91R-CC and 93R-CC through 96R-CC based on the occurrence of P. columnata in the absence of H. albiensis. Sample 210-1276A-97R-CC is barren.
Paleogene
Most of the 20 Paleogene core catcher samples investigated shipboard are barren of planktonic foraminifers or contain rare to few specimens (<50 tests), and only 2 of these core catcher samples contain age-diagnostic planktonic foraminiferal assemblages (Table T7). A number of thin sections of Paleogene sandstones provide additional age information based on planktonic foraminifers. A thin section from Sample 210-1276A-1W, 47-49 cm, indicates undifferentiated upper middle Eocene to lower upper Eocene Zones P12-P15 based on the presence of Turborotalia pomeroli. However, the presence of latest Eocene-early Oligocene age calcareous nannofossils and dinocysts in Core 210-1276A-1W suggests that the T. pomeroli and other planktonic foraminifers are reworked from older deposits. Undifferentiated uppermost lower Eocene to middle Eocene Zones P9-P12 are suggested by the presence of Acarinina bullbrooki, A. primitiva, Globigerinatheka cf. G. index, Truncorotaloides rohri, and Morozovella cf. M. spinulosa in thin sections from Samples 210-1276A-7R-2, 35-39 cm, and 7R-4, 94-98 cm.
Sample 210-1276A-8R-CC, located below the unconformity separating lithologic Units 1 and 2, contains rare and tiny planktonic foraminifers and is also assigned to undifferentiated uppermost lower Eocene to middle Eocene Zones P9-P12 based on the presence of A. bullbrooki. Sample 210-1276A-14R-CC is assigned to upper Paleocene Subzone P4c based on the co-occurrence of Globanomalina pseudomenardii, Morozovella aequa, and Igorina albeari. The barren Eocene and Paleocene samples reflect either dissolution of foraminiferal tests during settling through the lysocline and CCD or postdepositional loss due to acidic conditions in sediments rich in organic matter. The greatest concentrations of planktonic foraminifer tests occur in mudstone with coarse siltstone or very fine sandstone laminae, or they are in very fine to fine grainstone turbidites. Preservation is generally moderate, and whole specimens are filled with calcite. Reworked Late Cretaceous (Campanian-Maastrichtian) planktonic foraminifers are noted in Samples 210-1276A-9R-CC and 14R-CC.
Most of the grainstone thin sections from Core 210-1276A-15R to Section 21R-4 contain rare to abundant reworked Late Cretaceous planktonic foraminifers. A mid- to late Maastrichtian age for the reworked material is indicated by the presence of diverse species, including Abathomphalus mayaroensis, Racemiguembelina fructicosa, Contusotruncana contusa, Globotruncana stuarti, and G. conica. Sample 210-1276A-15R-1, 124-126 cm, is assigned to upper Paleocene Zone P4 based on the presence of the nominate taxon Globanomalina pseudomenardii. The presence of Morozovella conicotruncana in the thin section from Sample 210-1276A-17R-4, 98-99 cm, indicates lower upper Paleocene Zone P3. Samples 210-1276A-18R-6, 52-55 cm, and 18R-6, 88-90 cm, contain Parasubbotina pseudobulloides, which suggests undifferentiated lower Paleocene Zones P1-P2.
Sample 210-1276A-21R-4, 19-21 cm, lies 20-30 cm above the K/T boundary. This thin section contains common small planktonic foraminifers indicative of basal Paleocene Zone P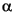 as well as the calcareous dinoflagellate Thoracosphaera spp. against a backdrop of abundant reworked upper Maastrichtian planktonic foraminifers of the Abathomphalus mayaroensis Zone (Zone KS31). Age-diagnostic taxa from this interval include Guembelitria cretacea, Parvularugoglobigerina eugubina, and Woodringina sp.
as well as the calcareous dinoflagellate Thoracosphaera spp. against a backdrop of abundant reworked upper Maastrichtian planktonic foraminifers of the Abathomphalus mayaroensis Zone (Zone KS31). Age-diagnostic taxa from this interval include Guembelitria cretacea, Parvularugoglobigerina eugubina, and Woodringina sp.
Upper Cretaceous
The richest assemblages of planktonic foraminifers are present in coarser-grained mudstones and very fine to fine sandstones that reflect downslope transport and size sorting. Sample 210-1276A-21R-CC is assigned to upper Maastrichtian Abathomphalus mayaroensis Zone (Zone KS31) based on the presence of the nominate taxon and a moderately diverse assemblage of globotruncanids and rugoglobigerinids (Table T7). A thin section from Sample 210-1276A-21R-4, 127-129 cm, contains a similarly diverse assemblage of planktonic foraminifers indicative of Zone KS31 including the taxa A. mayaroensis, Racemiguembelina fructicosa, and Contusotruncana contusa. Sample 210-1276A-22R-CC is missing species of the genus Gansserina but contains Globotruncana falsostuarti, which suggests the lower Maastrichtian part of Zone KS30. Mid-Maastrichtian marker taxa R. fructicosa and C. contusa are also absent from this core catcher sample. The absence or paucity of large biserial planktonic foraminifers (e.g., species of Racemiguembelina, Planoglobulina, and Gublerina), typical of Maastrichtian-age low-latitude (Tethyan) assemblages, suggests that Site 1276 may have been influenced by transitional or Boreal water masses at times during the latest Cretaceous. Alternatively, dissolution during settling may have selectively removed some of these larger taxa in all but the turbiditic sandstones. Sample 210-1276A-24R-CC is late Campanian in age (Zone KS28 or KS29) based on the presence of Globotruncana ventricosa and G. aegyptiaca and the absence of G. falsostuarti and Globotruncanita calcarata.
An elevated CCD (~2500-3000 m) in the North Atlantic Basin during the Aptian-Campanian (Thierstein, 1979; Tucholke and Vogt, 1979) exacerbated the problem of planktonic foraminiferal dissolution at the abyssal paleodepths of Site 1276. Samples 210-1276A-25R-CC through 28R-CC are barren of calcareous microfossils, a likely consequence of the high eustatic sea level, elevated CCD, and reduction of gravity flows to the deep sea during this part of the Cretaceous. Below this barren interval, a late early Turonian to early late Turonian age (Zone KS21) is indicated for Sample 210-1276A-29R-CC based on the co-occurrence of Marginotruncana renzi, M. sigali, Dicarinella imbricata, Praeglobotruncana praehelvetica, and Whiteinella aprica. However, planktonic foraminifers are very rare and poorly preserved in this sample.
Sample 210-1276A-30R-CC is barren of planktonic foraminifers, but thin sections from Samples 30R-5, 92-96 cm, and 31R-1, 81-84 cm, are assigned to uppermost Cenomanian-lowermost Turonian Whiteinella archaeocretacea Zone (Zone KS20) based on the presence of the nominate taxon together with Dicarinella cf. D. caniculata and Praeglobotruncana cf. P. praehelvetica. The upper Cenomanian Rotalipora cushmani Zone (Zone KS19) is recognized in the thin section from Sample 210-1276A-31R-5, 18-20 cm, based on the presence of R. cushmani and W. archaeocretacea.
Samples 210-1276A-31R-CC through 42R-CC are barren of planktonic foraminifers or contain only very tiny (<125 µm) specimens. The latter type of sample contains very fine to fine grains of quartz sand that are of similar size to the foraminifers, suggesting that the microfossils were size sorted during transport in distal turbidity currents. Despite the evidence for vigorous transport energy, these foraminiferal assemblages are typically moderately to moderately well preserved and contain species of the genera Heterohelix, Hedbergella, Globigerinelloides, Guembelitria, and Schackoina. Sample 210-1276A-35R-CC contains a moderately diverse assemblage that includes the species Hedbergella delrioensis, H. planispira, Globigerinelloides bentonensis, G. ultramicrus, Clavihedbergella simplex, and Praeglobotruncana delrioensis. However, the lack of any species of the genus Rotalipora (including juvenile specimens) in this sample may be another indication that cooler transitional or Boreal surface waters influenced the area of Site 1276 during the Cenomanian. A thin section from Sample 210-1276A-36R-2, 123-127 cm, is assigned to lower Cenomanian Rotalipora globotruncanoides Zone (Zone KS17) based on the presence of the nominate taxon together with Rotalipora appenninica, P. delrioensis, P. stephani, and Hedbergella libyca. The presence of the last taxon suggests that this sample lies in the basal Cenomanian because H. libyca is known to occur in a narrow zone straddling the Albian/Cenomanian boundary interval (Leckie, 1984).
Lower Cretaceous
The upper Albian at Site 1276 is characterized by moderately preserved and rare to abundant assemblages of planktonic foraminifers. These assemblages do not show the pronounced size sorting observed in the Cenomanian-Turonian interval. A thin section from Sample 210-1276A-41R-2, 88-91 cm, indicates upper Albian Rotalipora appenninica Zone (Zone KS16) based on the presence of R. appenninica in the absence of R. globotruncanoides. Samples 210-1276A-43R-CC and 44R-CC are likewise assigned to uppermost Albian Zone KS16 based on the presence of R. appenninica, R. ticinensis, Planomalina buxtorfi, Praehedbergella delrioensis, and Hedbergella libyca. Sample 210-1276A-45R-2, 112-114 cm, likely belongs to the same zone based on the co-occurrence of R. appenninica and P. buxtorfi. Upper Albian Zone KS15 occurs in Samples 210-1276A-45R-CC through 48R-CC. This zone is characterized by the presence of Rotalipora ticinensis, R. subticinensis, Ticinella roberti, T. primula, Biticinella breggiensis, and Globigerinelloides bentonensis. The greater abundance of planktonic foraminifers parallels generally higher concentrations of carbonate along continental margins during the late Albian, as well as larger sizes and more calcified species of planktonic foraminifers at this time (Leckie et al., 2002).
The interval from Samples 210-1276A-49R-CC to 80R-CC again contains sporadic occurrences of planktonic foraminifers, with a mix of either barren or size-sorted assemblages of juvenile specimens similar to those of the Cenomanian-Turonian interval. Samples 210-1276A-52R-CC and 54R-CC contain assemblages indicative of the lower part of upper Albian Zone KS14 based on the presence of Biticinella breggiensis, Ticinella praeticinensis, T. primula, and T. raynaudi. Sample 210-1276A-54R-CC, like Sample 48R-CC, is also an exception to typical mid-Cretaceous (Albian-Turonian) samples in that both samples contain abundant planktonic foraminifers in addition to few to abundant calcareous and agglutinated foraminifers and radiolarians. Sample 210-1276A-59R-CC contains T. primula and Hedbergella planispira in the absence of Biticinella and other species of Ticinella and indicates middle Albian Zone KS13. Samples 210-1276A-63R-CC and 73R-CC may also be assigned to Zone KS13, but the specimens are very small and too few in number to make a firm age determination. Most of the core catcher samples from Sections 210-1276A-74R-CC through 97R-CC are barren of planktonic foraminifers.
The utility of benthic foraminifers as stratigraphic index markers is limited by the long ranges and/or the geographic occurrence patterns of the majority of taxa. The group is most useful when considered in succession as characteristic assemblages, particularly in areas where regional stratigraphic zones have been proposed and tested (e.g., Kuhnt and Kaminski, 1997). Benthic foraminifers also are particularly useful as indicators of depositional environments.
Benthic foraminiferal assemblages recovered from Site 1276 fall into two associations that correspond to different stratigraphic intervals. Calcareous assemblages characterize the upper Eocene, lower Eocene to upper Paleocene, and Maastrichtian to upper Campanian. Agglutinated species dominate the assemblages in the remainder of the Eocene to Paleocene and in the upper Campanian to Albian sediments. The preservation of Paleogene and Maastrichtian calcareous benthic foraminifers is poor to moderate. Albian calcareous benthic foraminifers are well to moderately preserved in black shale and mudstone, and they are generally poorly preserved in sandstone lithofacies. Preservation of agglutinated foraminifers is variable among samples, and many taxa are commonly deformed. All tube-shaped forms such as Rhizammina, Bathysiphon, and Hyperammina are fragmented. Samples 210-1276A-3R-CC, 5R-CC, 20R-CC, 37R-CC, 51R-CC, 78R-CC through 80R-CC, 83R-CC, 85R-CC, 86R-CC, 89R-CC, 90R-CC, and 94R-CC through 96R-CC are barren. The distribution of benthic foraminifers is reported in Table T10.
Rare calcareous benthic foraminifers, including Bulimina tuxpamensis and Cibicidoides praemundulus, are present in the upper Eocene Sample 210-1276A-2R-CC. Agglutinated species are not observed in this sample. Eocene to Paleocene benthic foraminiferal assemblages fall into two categories that correspond to different lithologies. Assemblages in mudstone are typically characterized by various agglutinated species of the genera Ammodiscus, Spiroplectammina, Trochammina, and fragments of tube-shaped species of the genera Rhizammina and Bathysiphon (Samples 210-1276A-6R-CC, 7R-CC, and 15R-CC). Reticulophragmium sp. and Cyclammina cancellata are present in Samples 210-1276A-6R-CC and 7R-CC. The presence of these agglutinated taxa in the absence of Nuttallides truempyi indicate an abyssal paleodepth below the CCD (e.g., Kuhnt and Collins, 1996). The same constituents are observed in Samples 210-1276A-4R-CC, 10R-CC, 11R-CC, and 16R-CC through 19R-CC, although their abundances are very low. In contrast, sandy Samples 210-1276A-8R-CC, 9R-CC, 12R-CC, 13R-CC, and 14R-CC in lithologic Unit 2 are characterized by calcareous species including Cibicidoides spp., Astrononion novozealandicum, Nonionella sp., and species of Gyroidinoides. These species are poorly preserved. The genus Cibicidoides lives attached to seaweed or coarse-grained materials (Kitazato, 1994), and it is a common constituent in the neritic zone (Murray, 1991). The genera Astrononion and Nonionella are likewise characteristic of shelfal depths (Murray, 1991; Leckie and Olson, 2003). Hence, it is judged that these calcareous species are derived from a neritic environment (<200 m) and are displaced downslope by turbidity currents.
Maastrichtian to Campanian benthic foraminiferal assemblages are recognized in Samples 210-1276A-21R-CC, 22R-CC (claystone), 22R-CC (sandstone), and 24R-CC. These assemblages are characterized by moderately diverse calcareous species, including polymorphinids, Bolivina spp., buliminids, Gavelinella spp., and Gyroidina spp., whereas agglutinated forms are rarely present. These calcareous taxa are believed to be common constituents in bathyal (200-2000 m) to abyssal (>2000 m) depths (Sliter and Baker, 1972; Tjalsma and Lohmann, 1983). Aragonia velascoensis is an abyssal calcareous benthic foraminifer (Tjalsma and Lohmann, 1983) that also occurs in a sandstone in Sample 210-1276A-22R-CC. As such, these calcareous assemblages are interpreted to be a mixed fauna of in situ abyssal and transported bathyal taxa.
A sample examined from Section 210-1276A-23R-4 yields an assemblage of planktonic foraminifers together with common specimens of the agglutinated foraminiferal taxon Caudammina gigantea, a characteristic element of deepwater Upper Cretaceous sediments. The foraminiferal assemblage is moderately well preserved and not size sorted, which indicates that this sample contains in situ benthic foraminifers. This evidence suggests that the depositional environment was above the CCD (e.g., Kuhnt and Urquhart, 2001).
Campanian to Albian foraminiferal assemblages from mudstone are characterized by moderately diverse agglutinated forms, including Lagenammina spp., Glomospira spp., Ammobaculites spp., Ammodiscus spp., Spiroplectammina spectabilis, Trochammina spp., and fragments of tube-shaped species (Samples 210-1276A-24R-CC to 97R-CC). Similar taxa have been reported from the mid-Cretaceous at other abyssal sites in the North Atlantic (Gradstein, 1978; Sigal, 1979). Gyroidinoides cf. G. nitidus and Gavelinella cf. G. intermedia occur rarely but almost continuously between Samples 210-1276A-43R-CC and 74R-CC (upper to middle Albian). Because preservation of these calcareous species is moderate to good, it is possible that they lived together with agglutinated taxa in the abyssal zone above the depth of the paleo-CCD. However, the abundance of agglutinated species and the absence or paucity of calcareous bathyal forms suggests an abyssal paleodepth below the CCD. The presence of the genus Spiroplectammina is interesting, as this taxon incorporates calcium carbonate cement into the test wall. Thus, it is expected that these forms would not inhabit environments below the CCD. However, the possibility that these specimens of Spiroplectammina, like the calcareous benthic genera Gyroidinoides and Gavelinella, have been redeposited from upslope sites above the CCD and then subjected to rapid burial cannot be entirely discounted.
In contrast, benthic foraminifers are rarely observed in sandstone samples. However, small Praebulimina spp., Bolivina spp., and Gyroidina spp. occur sporadically (e.g., Samples 210-1276A-75R-CC and 76R-CC). Praebulimina spp. and Bolivina spp. are thought to be characteristic taxa of the oxygen minimum zone in upper bathyal depths (200-600 m) (Nyong and Olsson, 1984; Holbourn et al., 1999; Leckie and Olson, 2003). These calcareous forms are likely to have been transported from upper bathyal depths by turbidity currents.
Paleogene
Eleven of the twenty core catcher samples from the Paleogene that were investigated aboard the ship contain palynomorph assemblages with very good preservation. Lithologic Units 1 and 3 are represented in seven and four palynomorph-bearing samples, respectively. Core catcher samples from lithologic Unit 2 are barren of palynomorphs (Table T11). All Paleogene assemblages investigated are strongly dominated by dinoflagellate cysts (dinocysts). Other marine palynomorphs (acritarchs, prasinophytes, and foraminifer test linings) are generally present as single specimens only. Terrestrial palynomorphs, which are present in low abundances throughout the Paleogene intervals investigated, mainly comprise bisaccate pollen.
Sample 210-1276A-1W-CC is assigned an early Rupelian age (33.7-32 Ma; early Oligocene) based on the presence of Areoligera semicirculata and Glaphyrocysta semitecta. The maximum age of this sample has been determined based on the mid-Northern Hemisphere FO of 33.7 Ma for A. semicirculata (Williams et al., unpubl. data [N1]). Thus, this age assignment is more reliable than if it were based on LOs alone. This early Oligocene age assignment is supported by the absence of Lentinia serrata, which has a mid-Northern Hemisphere LO of 33.5 Ma (Williams et al., unpubl. data [N1]).
The dating of Sample 210-1276A-2R-CC as ~37.0 Ma (Priabonian to earliest Rupelian; late Eocene to earliest Oligocene) is based on the presence of Enneadocysta pectiniformis and Lentinia serrata. However, Areosphaeridium dictyoplokum, whose LO has been shown to be a reliable marker for the Eocene/Oligocene boundary in many Tethyan and northern European sections (e.g., Brinkhuis and Visscher, 1995), is only found lower in the section. Sample 210-1276A-3R-CC is dated as Priabonian (~37.0-33.6 Ma; late Eocene) based on the presence of Hemiplacophora semilunifera, E. pectiniformis, and L. serrata. Cerebrocysta bartonensis is present in this sample, in disagreement with the previously described range of this species in the northern mid-latitudes (41-38 Ma) (Williams et al., unpubl. data [N1]). It has yet to be resolved if this inconsistency is due to biogeographic differences or to reworking. The presence of Charlesdowniea coleothrypta and Dracodinium condylos is ascribed to reworking of Ypresian (lower Eocene) and Lutetian (lower middle Eocene) strata. Sample 210-1276A-4R-CC has been tentatively assigned an early Priabonian to late Bartonian age (latest middle Eocene; 38?-36.5 Ma) based on the presence of Adnatosphaeridium multispinosum, E. pectiniformis, and Phthanoperidinium echinatum. Other taxa typical for this interval are Charlesdowniea clathrata, Heteraulacacysta leptalea, Rhombodinium draco, and Wetzeliella spinula. Reworking is indicated by findings of Areoligera senonensis and Cerodinium wardense, both of which have Lutetian LOs.
Samples 210-1276A-5R-CC, 6R-CC, and 7R-CC contain Bartonian (upper middle Eocene) assemblages (40-38.5, 41-39, and 41-38 Ma, respectively). Age control is primarily based on the presence/absence of A. multispinosum, Cerebrocysta bartonensis, Diphyes colligerum, Lentinia serrata, Phthanoperidinium echinatum, and Rhombodinium draco. The presence of the Oligocene marker taxon Phthanoperidinium amoenum in Sample 210-1276A-5R-CC is attributed to downcore contamination resulting from drilling. For Sample 210-1276A-7R-CC, the presence of A. senonensis again indicates reworking of Lutetian (lower middle Eocene) or older strata.
Other than single reworked specimens identified in the Priabonian to Bartonian samples (see above), Selandian to Lutetian (lower upper Paleocene-lower middle Eocene) palynomorph assemblages were not observed at Site 1276. Samples 210-1276A-8R-CC through 15R-CC proved to be barren of palynomorphs. This is attributed to the strongly oxidized or the partly secondary, reduced nature of the respective sediments.
Assemblages suggesting Danian (early Paleocene) ages were identified in Samples 210-1276A-16R-CC to 18R-CC and 20R-CC. In this interval, palynological age control is predominantly based on the presence of Alisocysta circumtabulata, A. margarita, A. reticulata, Carpatella cornuta, Cerodinium diebelii, Damassadinium californicum, Oligosphaeridium pulcherrimum, Palaeocystodinium australinum, Palaeoperidinium pyrophorum, Senoniasphaera inornata, and Spongodinium delitiense. Species of the genus Alisocysta, which are among the most age-diagnostic taxa for the early Paleogene, are rare at Site 1276. The presence of Achomosphaera alcicornu in Sample 210-1276A-16R-CC, which predates the commonly used mid-northern latitude FO of this species by at least 4.2 m.y., may be due to either downcore contamination or an earlier appearance of A. alcicornu at Site 1276 than at more southerly locations.
Cretaceous
Nine of the twenty-one core catcher samples from the Cretaceous that were investigated aboard ship yield palynomorph assemblages with very good to excellent preservation. All productive samples are from lithologic Unit 5 (uppermost Aptian[?]-lower Albian to lowermost Turonian). Samples from the Cretaceous part of lithologic Unit 3 as well as from lithologic Unit 4 are barren of palynomorphs (Table T12). There is a downhole trend toward increasing terrestrial influence (documented in increasing amounts of terrigenous palynoclasts and sporomorphs) in lithologic Unit 5. Dinocysts are, however, also abundant in these samples.
Samples 210-1276A-29R-CC and 30R-CC are the youngest productive samples from the Cretaceous at Site 1276. They were assigned late Turonian (92.5-88.7 Ma) and early Turonian (93.5-92.5 Ma) ages, respectively, based on the presence of Chatangiella verrucosa, Epelidosphaeridia spinosa, Litosphaeridium siphoniphorum, Senoniasphaera rotundata alveoata, and Trithyrodinium suspectum. Extremely abundant terrestrial phytoclasts (mainly black wood) in Sample 210-1276A-29R-CC indicate a strong allochthonous signal that is fully compatible with the sedimentologically inferred turbiditic origin. Sample 210-1276A-31R-CC is dated as mid-Cenomanian to early Turonian (96-92.5 Ma) based on the LO of E. spinosa and the FO of Florentinia deanei. Other characteristic dinocyst taxa of this interval include Oligosphaeridium asterigum, Pervosphaeridium truncatum, and Tehamadinium mazaganensis.
Cenomanian ages were assigned to Samples 210-1276A-32R-CC, 33R-CC, and 39R-CC (100.5-92.5, 100-95.8, and 98.9-96 Ma, respectively). Palynological age control in the Cenomanian of Site 1276 is mainly based on the presence of Cyclonephelium membraniphorum, Epelidosphaeridia spinosa, Hapsocysta peridictya, Litosphaeridium siphoniphorum, Ovoidinium verrucosum, and Xiphophorum alatum. All three samples exhibit high abundances of Palaeohystrichophora infusorioides.
Dinocyst assemblages from Samples 210-1276A-41R-CC and 45R-CC yield late Albian ages (100-98.9 and 103.2-101.06 Ma, respectively). Diagnostic taxa are, among others, Carpodinium obliquicostatum, Chichaodinium vestitum, Litosphaeridium siphoniphorum, L. conispinum, and Ovoidinium verrucosum. Sample 210-1276A-56R-CC is assigned a middle Albian age (107-103 Ma) based on the presence of Callaiosphaeridium cf. C. trycherium, Carpodinium granulatum, Dinopterygium cladoides, and Stephodinium coronatum.
The evidence presented above indicates very good preservation of palynomorphs in combination with high sedimentation rates. Thus, there is excellent potential for in-depth palynological studies covering the interval from the Turonian through the Albian in Site 1276 sediments.
Radiolarians have previously been recorded in abundance from sediments deposited in the North Atlantic during Paleogene to Cretaceous time. The most notable radiolarian blooms previously documented in the literature are recorded in sediments deposited during early Eocene and Campanian times and in the Albian/Cenomanian boundary interval (Thurow, 1988). Radiolarian faunas are also a common component of Mesozoic Tethyan sediments. As such, it was anticipated that these chronological intervals would prove to be the most productive for radiolarian recovery during Leg 210. Mesozoic radiolarians are usually preserved as opal-CT, quartz, or pyrite, depending on diagenetic history. Mineral replacement of the test does not commonly obscure identifying features of the taxa, and in some instances the diagenetic replacement can be advantageous to fossil preservation. Palynological preparations processed from Site 1276 yielded pyritized fragments of both radiolarians and diatoms, giving a good indication that the depositional environments were normal open-marine conditions. The presence of these fragments helps us to focus on the potentially most productive samples for further, more rigorous processing methods.
At Site 1276, Paleogene radiolarian faunas were recovered from the >63-µm fraction of the washed residues of Samples 210-1276A-5R-CC, 9R-CC, 10R-CC, 11R-CC, and 13R-CC. The assigned calcareous nannofossil age for these samples is early to middle Eocene, consistent with widespread siliceous deposits of Eocene age in the Atlantic Ocean. Radiolarians were also recovered from Samples 210-1276A-14R-CC and 18R-CC, which are Paleocene in age based on calcareous nannofossils. Although all these Paleogene occurrences are sometimes abundant, the preservation of the specimens is too poor for positive identification. The assemblages consist mostly of small and seemingly spherical forms.
Published occurrences of Mesozoic radiolarians from North Atlantic sediments recorded to date are few, and the most notable descriptions include those by Foreman (1977), Empson-Morin (1984), and Thurow (1988). In the Mesozoic section cored at Site 1276, rich and diverse radiolarian assemblages were recovered from only three core catcher samples: 210-1276A-48R-CC, 54R-CC, and 96R-CC. Samples 210-1276A-48R-CC and 54R-CC are upper Albian sediments of lithologic Subunit 5B, a lithologic unit characterized by dark greenish gray, medium brown, and greenish black mudstones and claystones. In both of these samples, the radiolarian assemblages are associated with abundant planktonic foraminifers. The radiolarian tests occur as opal-CT and are moderately well preserved. The diverse assemblages are dominated by nassellarians including abundant cryptocephalic taxa together with species of Xitus, Archaeodictyomitra, Dictyomitra, and Amphipyndax. The assemblages differ slightly from typical Albian-age Tethyan faunas in that there are very few spumellarians. In particular, the characteristic Mesozoic pseudoaulophacid and hagiastrid taxa are absent. The assemblages show components and preservational characteristics similar to radiolarian faunas recovered from the Albian Gault Clay of southern Britain (Urquhart and Wilkinson, 1997). The radiolarian assemblage recovered from Sample 210-1276A-96R-CC is quite different in terms of preservation in that the majority of specimens are replaced by pyrite. This replacement mineralization has not, however, obscured the external morphologies used for identification of Mesozoic radiolarians. The specimens are moderately well preserved, and the assemblage has a typically Aptian-Albian faunal aspect. Despite this, the sample has not yet yielded age-diagnostic marker species, the presence of which would allow a more refined zonal assignment to be placed on this sample.
Small (<125 µm) pyritized and indeterminate centric diatoms are common in the >63-µm size fraction of the washed residue of the lower Paleocene dark gray mudstone in Sample 210-1276A-20R-CC. As mentioned above, pyritized diatoms and diatom fragments are also present in many of the palynological preparations.
Ichthyoliths (fish teeth) and fish bones are present in most samples throughout the hole but are rare; this suggests normal marine conditions but poor preservational environments. Ostracodes are recovered sporadically in the mid-Albian section in low abundances, usually as single specimens. Calcite prisms of inoceramid bivalve shells are present in a number of Albian-age samples. These prisms are a common and characteristic component of Cretaceous sediments, especially those deposited during the Campanian and early Maastrichtian. Their relative scarcity in sediments recovered at Site 1276 suggests depositional environments below the CCD.
Sedimentation rates were determined by connecting straight-line segments through median depths in an age-depth plot depicting key calcareous nannofossil FOs or LOs (Fig. F141). Select palynomorph FOs and LOs are also shown. In Figure F142, the minimum and maximum ages of calcareous nannofossil, planktonic foraminifer, and palynomorph zones (or ages) for each sample are shown by conjugate triangles that are color-coded by microfossil group. See Table T12 for the datums used to construct the age-depth curve for Site 1276 and Table T13 for the datums used to construct the age-depth curve for DSDP Site 398.
The first core taken at Hole 1276A is a wash core (210-1276A-1W), and its placement in the interval from ~753 to ~800 mbsf is uncertain; therefore, biostratigraphic datums from Core 1W are not used in the sedimentation rate calculations. The age-depth plot reveals a number of marked changes in sedimentation rate; at least one, and possibly two, unconformities; and two condensed intervals. The changes in slope correspond closely to lithologic unit boundaries (see "Lithostratigraphy"). Samples 210-1276A-2R-CC through 7R-CC (basal Oligocene to middle Eocene) correspond to lithologic Unit 1 and show an average sedimentation rate of ~7.4 m/m.y. based on calcareous nannofossil datums and a range of 5.7-9.7 m/m.y. based on palynomorphs.
The apparent discrepancy between the (younger) palynomorph and (older) calcareous nannofossil-based ages in Sections 210-1276A-2R-CC through 7R-CC can be explained in several ways, the foremost being dissolution of calcareous microfossils in combination with extensive reworking. Deposition of sediments below the CCD probably reduced the numbers of in situ calcareous nannofossils that were then diluted by allochthonous (reworked) specimens. The dissolution effect can be excluded for the (organic walled) palynomorphs. Moreover, the palynomorph data for this time interval rely more on FOs in comparison to the nannoplankton data, which makes them less prone to problems associated with reworking. Another factor contributing to the age discrepancies between the calcareous nannofossil and dinoflagellates in the middle Eocene-lower Oligocene of Hole 1276A was the use of a low-latitude tropical calcareous nannofossil zonation for a site that was, at times, influenced by cooler-water masses (the paucity of Discoasters at this site could be evidence of this influence); therefore, biostratigraphic data based on Discoasters may be less reliable. In addition, direct calibration of dinoflagellate events with magnetic polarity stratigraphy in deep-ocean cores is in its infancy.
Calcareous nannofossil datums in the upper part of Core 210-1276A-2R suggest the presence of an unconformity or condensed interval spanning the upper middle and upper Eocene, although additional shore-based research is required to constrain the duration. A second, deeper unconformity spanning as much as 8.5 m.y. may be present at the boundary between lithologic Units 1 and 2 in Section 210-1276A-8R-5. This estimate is based on the combination of young FOs of the dinocysts Lentinia serrata and Rhombodinium draco in Sample 210-1276A-6R-CC and the much older FO of the calcareous nannoplankton taxa Rhabdosphaera inflata in Sample 8R-CC. An erosional event of much shorter duration would be indicated by the calcareous nannoplankton datums alone, but barren samples in Cores 210-1276A-4R through 7R hampered shipboard resolution of the unconformity.
An average sedimentation rate of 7.4 m/m.y. persists through the middle and lower Eocene (Samples 210-1276A-8R-CC to 14R-1, 104 cm) of Site 1276 based on the calcareous nannofossil data. This interval corresponds to much of lithologic Unit 2. The Paleocene-upper Maastrichtian interval corresponds to the basal part of lithologic Unit 2 and much of lithologic Unit 3, and it accumulated at an average rate of ~6.4 m/m.y. The lower Maastrichtian is condensed or missing at this site and is represented by the red sediments of Core 210-1276A-22R. The calcareous nannofossil datums found in the lower part of lithologic Unit 3 suggest that the upper Campanian sediments accumulated at an average rate of ~4.3 m/m.y.
An interval of paleontologically barren samples occurs in Cores 210-1276A-25R through 28R. This interval represents the bulk of lithologic Unit 4, which accumulated at an average rate of ~1.9 m/m.y. based on the bracketing calcareous nannofossil datums above and below. Because of the lack of age control, this interval could include an undetected hiatus. The Turonian (basal part of lithologic Unit 4 and uppermost part of Unit 5) accumulated at an average rate of 4.0 m/m.y. The middle Albian-Cenomanian part of lithologic Unit 5 (Cores 210-1276A-31R through 51R) accumulated at an average sedimentation rate of 17.8 m/m.y., whereas the middle Albian (Cores 52R-77R) accumulated at a greatly increased average rate of ~104.9 m/m.y. The uppermost Aptian(?)-lower Albian interval (Cores 210-1276A-78R through 97R) records an average sedimentation rate of 22.3 m/m.y. The overall pattern of changing sedimentation rates at Site 1276 is strikingly similar to that of DSDP Site 398 on the conjugate margin of the Iberia Peninsula (Figs. F141, F142).
