ACCOMPLISHMENTS
Evaluated Sequence Variability
We confirmed the conclusions of Legs 150 and 150X (see summary in Miller et al., 1997) that eustasy is the dominant process that determines the template of potential sequences and their general architecture (stacking patterns and preservation of stratal surfaces). We attribute some of the differential preservation of sequences in boreholes from DE through NJ to minor (~10-m scale) tectonic differences caused by minor movement on basement blocks (e.g., the rolling basins concept of Owens et al., 1997). Sediment supply can also locally affect the preservation of sequences, with preferential preservation of sequences near depocenters resulting from excess accommodation by loading and preferential removal in regions of sediment starvation. Sediment supply also determines the paloeoenvironmental setting (e.g., wave vs. deltaic dominated) (Fig. F2), the facies expressed regionally and locally, and the local expression of stratal surfaces.
Sequence Correlation with Global Records
We identified, dated, and correlated sequences among sites in NJ and DE with other global records (Fig. F3) (e.g.,  18O), confirming the link between sequence boundaries and 18O determined by Miller et al. (1996b) based on Leg 150 and 150X studies. Sequence boundaries are unconformities that are recognized on the basis of physical stratigraphy, including irregular contacts, reworking, bioturbation, major facies changes, gamma ray peaks, and paraconformities inferred from age breaks. Sr isotopic stratigraphy provided the primary age control on Oligocene-Miocene sections supplemented by calcareous nannofossil, planktonic foraminiferal, and dinocyst biostratigraphy. During Leg 174AX, thick Miocene sequences at Ocean View and Bethany Beach were sampled, identified, and dated. It extended our record of sequences dated onshore from ~10 Ma (Leg 150X) to 8 Ma, complementing Leg 174A drilling.
18O), confirming the link between sequence boundaries and 18O determined by Miller et al. (1996b) based on Leg 150 and 150X studies. Sequence boundaries are unconformities that are recognized on the basis of physical stratigraphy, including irregular contacts, reworking, bioturbation, major facies changes, gamma ray peaks, and paraconformities inferred from age breaks. Sr isotopic stratigraphy provided the primary age control on Oligocene-Miocene sections supplemented by calcareous nannofossil, planktonic foraminiferal, and dinocyst biostratigraphy. During Leg 174AX, thick Miocene sequences at Ocean View and Bethany Beach were sampled, identified, and dated. It extended our record of sequences dated onshore from ~10 Ma (Leg 150X) to 8 Ma, complementing Leg 174A drilling.
Facies Models
We developed facies models for deltaic-dominated (NJ) (Fig. F2A) and wave-dominated (DE) (Fig. F2B) successions. The deltaically influenced Miocene sections in NJ fit a facies model similar to the Niger Delta (Fig.
F2A) (Allen, 1970). Facies include marsh deposits, nearshore to delta front sands, prodelta silty clays, and shelfal sands (glauconitic in Paleogene and older sections and primarily quartz in the Miocene) (Fig.
F2A) (Allen, 1970). The facies are arranged as basal transgressive sands, medial prodelta silty clays, and upper quartz sands. The DE Miocene and younger sequences sampled at Bethany Beach consist of silts and sands deposited in fluvial to upper estuarine, lower estuarine, upper shoreface/foreshore, distal upper shoreface, lower shoreface, and inner and middle neritic environments (Fig.
F2B).
Intrasequence Architecture
We evaluated intrasequence architecture. Despite fundamentally different sedimentary regimes (wave-dominated shorelines in DE vs. deltaic systems in NJ), both regions share a similar sequence stratigraphic signature for the Miocene. LSTs are largely absent, and thus transgressive surfaces are usually merged with sequence boundaries; the exceptions include lowstand deposits identified for the first time in the Miocene Kirkwood Formation at Ocean View in the Kw2a and Kw1a sequences. TSTs are present at the bases of some sequences but are thin. In general, HSTs are divided into a lower fine-grained unit (silty clay in NJ and generally silts in DE) and an upper sandy unit. The upper HST sands comprise important aquifers in both regions that are generally confined by the overlying lower HST. Aside from these similarities, there are important sequence stratigraphic differences between regions. Maximum flooding surfaces (MFSs) identified in the Bethany Beach borehole show much greater evidence of erosion than MFSs in NJ, whereas sequence boundaries are often more subtle in DE because of the juxtaposition of similar facies.
Higher-Order Variability
We discovered higher-order (400?-k.y. scale) variability within lower Miocene sequences at Ocean View and Bethany Beach embedded within the million-year-scale sequences defined onshore during Leg 150X drilling. The Kw2a and Kw1a sequences are each provisionally subdivided into three higher-order sequences (Kw2a1, Kw2a2, and Kw2a3 and Kw1a1, Kw1a2, and Kw1a3, respectively). The preservation of higher-order lower-middle Miocene sequences may be the result of higher sediment supply and accommodation space because they are only recognizable in sections with high (~100 m/m.y.) sedimentation rates. Alternatively, the great thickness of the section is consistent with an autocyclical cause (lobe switching) for these apparent sequences. Analyses of nearshore seismic profiles recently collected near Ocean View (Fig. F1) should reveal if the higher-order cyclicity is due to base level lowering and, hence, if these are eustatically controlled sequences.
Comparison of NJ and DE Sequences
Comparison of NJ and DE (Bethany Beach) Miocene sequences highlights regional differences in sedimentation and possible tectonic controls. Equivalents of the Kw3, Kw2c, Kw2b, Kw1c, and Kw1a NJ sequences are represented at Bethany Beach (Fig. F3), although the sequences are generally thicker and sedimentation rates are higher in DE. Nevertheless, thickness does not equate to stratigraphic continuity: the NJ record is much more complete in the early part of the early Miocene (19-23.8 Ma), with the Kw1b and Kw0 sequence apparently missing in DE. The DE section is more complete in the late part of the early Miocene (~19-16.2 Ma), with one sequence (18.0-18.4 Ma) not represented in NJ. The upper part of the Oligocene (~27-23.8 Ma; sequences O6 and O5 of Pekar et al., 2000) is also absent at Bethany Beach because of truncation. These minor differences in preservation may be due to (1) minor tectonic movements (tens of meters) of basement blocks (Owens et al., 1997) or (2) differential preservation resulting from local sediment loading.
Oligocene Strata Geometry Reconstruction
The geometry of Oligocene strata in NJ was reconstructed using two-dimensional flexural backstripping (Pekar et al., 2000; Kominz and Pekar, 2001), and water depths were estimated for the margin using two-dimensional paleoslope modeling of the benthic foraminiferal biofacies (Pekar and Kominz, 2001; Pekar et al., in press b). These results allowed quantitative evaluation of factors controlling sedimentation on passive margins and comparison with other stratigraphic models (Fig. F2C) (Pekar et al., in press b). NJ Oligocene sequences are highstand dominated, TSTs are thin, and LSTs are absent. Pekar et al. (in press b) suggested that the absence of LSTs was due to bypassing because of efficient transfer of sediment across the shallow shelf, combined with the absence of major river systems in the area of study. They concluded that well-developed offlap at each sequence boundary is due primarily to marine bypassing and degradation, rather than to "forced regression" (Pekar et al., in press b).
Stratigraphic Response to Eustatic Change
Stratigraphic architecture observed at the NJ continental margin was also compared with Oligocene eustatic records to evaluate the stratigraphic response to eustatic change (Pekar et al., 2001). Early to "mid"-Oligocene sequence boundaries (33.8-28.0 Ma) were associated with relatively long hiatuses (0.3-0.6 m.y.), in which sedimentation in many places terminated during eustatic falls and resumed early during eustatic rises. Late Oligocene sequence boundaries were associated with relatively short hiatuses (<0.3 m.y.); they provide the best constraints on phase relations between sea-level forcing and margin response. The interval represented by each late Oligocene sequence varies in dip profile. At updip locations landward of the clinoform rollover in the underlying sequence boundary, sedimentation commenced after the eustatic low and terminated before the eustatic high (with partial erosion of any younger record) (Fig.
F2C). At downdip locations, sedimentation within each sequence was progressively delayed in a seaward direction, beginning during the eustatic rise and terminating near the eustatic low (Fig.
F2C). Combining data from all available boreholes, Pekar et al. (2001) showed that the ages of sequence boundaries (correlative surfaces) correspond closely to the timing of eustatic lows, and ages of condensed sections (intervals of sediment starvation) to eustatic highs (Fig.
F2C).
Core-Log Integration
Core log-downhole log integration provided a means of further evaluating sequences and variability within sequences. Lanci et al. (2002) integrated core-log magnetic susceptibility (MS) and natural gamma ray (NGR) measurements from Ancora with lithology and the downhole NGR log. They used a simple linear model to explain MS and NGR values by lithologic variation. Spectral NGR shows that high gamma values are due to the 40K radioisotope associated with glauconite and clay, sediment components that also tend to give high MS values. The major deviation from their model was at an anomalous level with high NGR but low MS values; this level was attributed to high uranium concentration in phosphorite. With only few exceptions, sequence boundaries identified at Ancora are expressed in the NGR and/or MS logs.
Backstripping and Eustatic Estimates
Leg 174AX provided material suitable for one- and two-dimensional backstripping of mid-Cretaceous to Miocene sections, providing a eustatic estimate for the interval from 100 to 8 Ma. Backstripping analysis of the Bass River and Ancora boreholes (Fig. F4) (Van Sickel et al., unpubl. data) provides complete Late Cretaceous sea-level estimates and tests previously published Cenozoic sea-level estimates based on Leg 150X drilling (Fig. F4)
(Kominz et al., 1998). Van Sickel et al. (unpubl. data) used electric logs to provide a new porosity-depth calibration for decompacting sediments, showing considerably lower porosity than those previously calculated at the offshore Cost B-2 well. Amplitudes and duration of sea-level changes were comparable when sequences were represented at multiple borehole sites, suggesting that the resultant curves are an approximation of regional sea level (Fig. F4). Sea-level amplitudes as great as 50 m are associated with Cretaceous sequences, whereas most Late Cenozoic amplitudes were closer to 20 to 40 m (Fig. F4) (Van Sickel et al., unpubl. data).
Two-dimensional backstripping of prograding Oligocene sequences reconstructed by Pekar et al. (2000) provided a detailed and precise eustatic estimate (Kominz and Pekar, 2001). Ten latest Eocene to earliest Miocene sequences provided the basis for estimates of ~20- to 60-m eustatic lowerings (Kominz and Pekar, 2001). The slightly higher eustatic estimates obtained by Kominz and Pekar (2001) vs. Van Sickel et al. (unpubl. data), particularly for earliest Oligocene rises and fall (see below) are attributed to the greater precision provided by two-dimensional backstripping that more nearly captured the full amplitude of eustatic change (Pekar and Kominz, 2001). Both one- and two-dimensional backstripping yielded Oligocene eustatic estimates that are lower than those published by the EPR group (e.g., Haq et al., 1987) by a factor of two or more (Kominz and Pekar, 2001; Van Sickel et al., unpubl. data). Oligocene eustatic lowerings were linked to global 18O increases (Pekar et al., 2001) and used to provide a sea level/18O calibration for the Oligocene (Pekar et al., in press a).
The Bass River and Ancora Cretaceous sections provided the means to estimate global sea-level (eustatic) variations of the Late Cretaceous (99-65 Ma) greenhouse world (Miller et al., unpubl. data). These two sites recorded 11-14 Upper Cretaceous sequences that were dated by integrating Sr isotopic stratigraphy and biostratigraphy (Fig. F5). The ages of sequence boundaries not only correlate regionally between sites (Fig. F5), they also correlate with the sea-level lowerings of EPR (Haq et al., 1987), northwest European (Hancock, 1993) and Russian sections (Fig. F6) (Sahagian et al., 1996), indicating a global cause. Backstripping yielded a Late Cretaceous eustatic estimate for these sequences, taking into account sediment loading, compaction, paleowater depth, and basin subsidence (Fig. F6). Sea-level changes were large (>25 m) and rapid (<1 m.y.), strongly suggesting glacioeustatic control of sea-level variations during the Late Cretaceous. Though the timing of EPR eustatic lowerings may be more or less correct, the EPR curve cannot be used as a valid Late Cretaceous eustatic record. Eustatic estimates from NJ and the Russian platform clearly show that the amplitudes of the major EPR eustatic lowerings were too high by a factor of at least two (Fig. F6). In addition, the EPR record differs in shape from the backstripped eustatic estimates. For example, the extremely large mid-Turonian and mid-Maastrichtian events reported by EPR are much lower in amplitude in the backstripped records, whereas the major flooding events at 69, 76, and 84 Ma in the NJ record are less important in the EPR record (Fig. F6).
Oxygen isotopic comparisons with Late Cretaceous sequence boundaries have not attained the resolution needed to unequivocally link the two as has been done for the past 42 m.y. (Miller et al., 1998a). Nevertheless, comparisons between Late Cretaceous sequence stratigraphy and 18O records are intriguing (Fig. F6) (Miller et al., unpubl. data), further suggesting small ice sheets in this alleged greenhouse world: (1) a major mid-Cenomanian sequence boundary (see also Gale et al., 2002) between the Potomac and Bass River I sequences (hiatus = ~96-97 Ma) correlates with a major (>1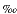 ) 18O increase; (2) two minor 18O increases spanning the Cenomanian/Turonian boundary may correlate with sequence boundaries at the base of Bass River II and Bass River III; and (3) a mid-Turonian sea-level lowering associated with the Bass River III/Magothy contact (91.5-92 Ma) may correlate with a major increase in benthic foraminiferal 18O values (~1.0), though additional data are needed to determine the precise timing of the increase (Fig. F6). Several other Coniacian-Campanian 18O increases (dashed arrows in Fig. F6) may be related to sequence boundaries, but the data are too sparse to provide a firm correlation. Miller et al. (unpubl. data) note that 18O data are consistent with a glacioeustatic cause for Late Cretaceous sequence boundaries. The data shown on Fig. F6 require that either large, rapid sea-level variations occurred during the Late Cretaceous greenhouse world or our understanding of causal mechanisms for global sea-level change is fundamentally flawed.
) 18O increase; (2) two minor 18O increases spanning the Cenomanian/Turonian boundary may correlate with sequence boundaries at the base of Bass River II and Bass River III; and (3) a mid-Turonian sea-level lowering associated with the Bass River III/Magothy contact (91.5-92 Ma) may correlate with a major increase in benthic foraminiferal 18O values (~1.0), though additional data are needed to determine the precise timing of the increase (Fig. F6). Several other Coniacian-Campanian 18O increases (dashed arrows in Fig. F6) may be related to sequence boundaries, but the data are too sparse to provide a firm correlation. Miller et al. (unpubl. data) note that 18O data are consistent with a glacioeustatic cause for Late Cretaceous sequence boundaries. The data shown on Fig. F6 require that either large, rapid sea-level variations occurred during the Late Cretaceous greenhouse world or our understanding of causal mechanisms for global sea-level change is fundamentally flawed.
Global Events in Earth History
Cenomanian/Turonian Boundary Events
Uppermost Cenomanian to lower Turonian strata are characterized by worldwide organic carbon-rich deposits (Ocean Anoxic Event 2 [OAE2]) (Arthur et al., 1985, 1987) reflected in a pronounced positive excursion of 13C (e.g., Schlanger et al., 1987; Jenkyns et al., 1994). The Bass River borehole recovered a thick (61 m) record of the latest Cenomanian to early Turonian that is continuous as measured by biostratigraphy (Sugarman et al., 1999) and cyclostratigraphy (Cramer in Wright et al., unpubl. data). Benthic foraminiferal 13C records show a large (>2) increase immediately below the C/T boundary (Fig. F7) (Sugarman et al., 1999). Above the sharp 13C increase, elevated 13C and sedimentary organic carbon (>0.9%) values continue into the lower Turonian, culminating in a sharp 13C decrease (Fig. F7) (Sugarman et al., 1999). High 13C values in the uppermost Cenomanian-lower Turonian at Bass River correlate with the OAE2, a global carbon burial event recorded in Europe and the U.S. western interior (Sugarman et al., 1999); we estimate that the duration of this event at Bass River is 850 k.y. based on cyclostratigraphy (Cramer in Wright et al., unpubl. data).
The OAE2 event occurred during long-term eustatic rise (10-m.y. scale), yet it occurs within a 1- to 2-m.y.-long sequence at Bass River and is not associated with maximum flooding. Thus, there is no relationship between OAE2 and sea-level lowering on the million-year scale (Fig. F7) (Sugarman et al., 1999). Within the sequence spanning the carbon event, there are at least four shallowing-upward parasequences (durations = ~350-460 k.y.) indicated by changes in abundance and type of Epistomina species, 18O variations, and minor lithologic variations (Fig. F7). There is no clear association between parasequence boundaries (Fig. F7) and OAE2, indicating that there is no relationship of the event and sea-level change on the 100-k.y. scale (Fig. F7) (Sugarman et al., 1999). Thus, Sugarman et al. (1999) concluded that whereas the organic carbon burial event was associated with a general long-term (10-m.y. scale) eustatic rise, the initiation and termination of the peak organic burial event itself were unrelated to sea-level change (Fig. F7).
Campanian/Maastrichtian Boundary Events
A ~71-Ma sequence boundary separating the base of the Maastrichtian Navesink Formation from the underlying Campanian Mount Laurel Formation (Fig. F5) is one of the most dramatic of the ~30 Cenozoic and 11-14 Late Cretaceous sequence boundaries identified in the NJ Coastal Plain (Miller et al., 1998a; Miller et al., 1999). This unconformity is found throughout the eastern U.S. (Owens and Gohn, 1985). The amount of eustatic lowering associated with this event is uncertain because of a hiatus, but ~30 m of eustatic rise is recorded at Ancora and Bass River, providing a minimum range of sea-level change (Fig. F6). The event has been linked to a 18O increase that occurred in both deep-sea benthic and low-latitude planktonic foraminifers (Fig. F6) (Miller et al., 1999). Based on this correspondence, Miller et al. (1999) argued that this event resulted from the growth of a transient ice cap (equivalent to ~40% the volume of the present-day east Antarctic ice cap) that caused a ~25- to 30-m glacioeustatic lowering at 71 Ma. The ~71-Ma Campanian/Maastrichtian boundary sea-level lowering was associated with a major reorganization in deepwater circulation as cool water from a high-latitude source influenced intermediate depths in the tropical Pacific (Barrera et al., 1997). A global 13C decrease is also correlated with the sea-level lowering, perhaps because of increased weathering of organic-rich sediments exposed on continental shelves (Barrera et al., 1997).
Latest Maastrichtian Events
Global 18O records show general cooling conditions in deep waters and high latitudes from the late Campanian to latest Maastrichtian (~73-66 Ma) (Fig. F6) (Barrera and Savin, 1999), punctuated by the ~71-Ma ice volume and sharp cooling event (Miller et al., 1999). During the latest Maastrichtian, planktonic foraminiferal distributions (particularly the thermophilic taxon Pseudotextularia elegans) and global 18O records (Figs. F6, F8) (Barrera and Savin, 1999) show a warming of sea-surface temperatures of ~5°C (Fig. F8) (Olsson et al., 2001). Planktonic foraminiferal 18O records from Bass River display this warming as a very large decrease that began at ~500 k.y. and ended about 22 k.y. before the K/T boundary (Fig. F8) (Olsson et al., 2001). Neritic benthic foraminiferal 18O records from Bass River show only a 0.2-0.5 coeval decrease (a 1°-2°C warming), indicating a strengthening in the thermocline at this time (Olsson et al., 2001). The latest Maastrichtian warming event may have been caused by a greenhouse effect resulting from the main outpouring of the Deccan Traps in India beginning at ~65.6 Ma (Courtillot et al., 1986). Sr isotopic data from Bass River (Fig. F8) (Olsson et al., 2002) and other sections (Vonhof and Smit, 1997) that show a distinct decrease from ~65.5 to 65 Ma are consistent with increased basaltic weathering. This warming may have contributed to the decline in dinosaur diversity (Sloan et al., 1986), though it was the impact at the K/T boundary that is unequivocally linked to the mass extinction of marine and presumably terrestrial taxa.
K/T Boundary Impact and Tsunami
The Bass River borehole provides a continuous depositional record across the K/T event (Olsson et al., 1997), which has been attributed to a bolide impact (Alvarez et al., 1980) near Chicxulub, Mexico (Hildebrand et al., 1991). K/T boundary sections in the Gulf Coast show thick ejecta, including glass spherules and tsunamites (Smit et al., 1992). Nonetheless, the relationship between ejecta and the marine mass extinction has been controversial (e.g., Keller et al., 1994), in part because the extensive mixing by a megatsunami in the Gulf Coast. In contrast, the U.S. East Coast was sheltered from the megatsunami's direct effects. The Bass River borehole directly ties impact ejecta to the highest occurrence of planktonic foraminifers and calcareous nannofossils (Fig. F9), providing the first unequivocal link between bolide impact and marine mass extinctions that mark the end of the Cretaceous (Olsson et al., 1997). Similar results were noted during ODP Leg 171B on the Blake Nose (Norris et al., 1999), with the ejecta present at the level of the marine mass extinction. A tsunami immediately followed the fallout of tektites from the Chicxulub ejecta vapor cloud in the Gulf of Mexico (Smit et al., 1992); a clast unit present above the K/T ejecta at Bass River (Fig. F9) (Olsson et al., 1997) and on the Blake Nose (Norris et al., 1999) also appears to be related to a tsunami. Although direct effects of the impact can explain tsunamites in the Gulf of Mexico, it has been difficult to explain the cause of tsunamites outside this region because the Florida platform would have attenuated the waves. Massive slumping on the Atlantic slope, including NJ, may have triggered a tsunami on the Atlantic margin as exemplified by a clast layer found at Bass River (Fig. F9) (Olsson et al., 1997, 2002), Ancora, and outcrop sections in the NJ Coastal Plain.
Though sea-level change has long been associated with the K/T boundary, the records from Bass River and Ancora clearly show that the impact event occurred within a sequence that spans the K/T boundary (the Navesink sequence; ~69-64.5 Ma) (Fig. F6) (Olsson et al., 2002). The sequence shallows upsection, particularly in the last 0.5 m.y. of the Cretaceous, culminating in a sharp shallowing in the last 100 k.y. of the Cretaceous (Fig. F8). However, there is no sequence boundary associated with the K/T boundary; a major sequence boundary is present in Biochron P1b, with a hiatus from ~64 to 63 Ma (Figs. F5, F6). Thus, Olsson et al. (2002) concluded that there was a minimal change in sea level associated with the K/T boundary.
Zachos and Arthur (1986) showed that the vertical carbon isotopic difference in the oceans (viz. between surface-dwelling planktonic and deep-sea [particularly Pacific] benthic foraminifers) disappeared at the K/T boundary. This "Strangelove" ocean was one with minimal export productivity (D'Hondt et al., 1998). Planktonic (Rugoglobigerina) and benthic (Anomalinoides) 13C records from Bass River (Fig. F8) (Olsson et al., 2002) demonstrate that the collapse of the vertical gradient also occurred in neritic sections. Sedimentation in the earliest Paleocene in the Mid-Atlantic shelf was dominated by slow glauconitic sedimentation, as both siliciclastic and pelagic input were low.
P/E Boundary Event
First recognized by Kennett and Stott (1990), the PETM is associated with a large (2.5-4), rapid (<20 k.y.) transient global carbon isotopic excursion (the CIE of Zachos et al., 1993), a negative excursion (1-3) in oxygen isotopes (e.g., Kennett and Stott, 1991), a deep-sea benthic foraminiferal extinction event (e.g., Tjalsma and Lohmann, 1983), a terrestrial mammalian turnover (e.g., Gingerich, 1989), and a kaolinite spike (Robert and Kennett, 1994). The CIE has been attributed to methane release from gas hydrate reservoirs (e.g., Dickens et al., 1995; Katz et al., 1999). Gibson et al. (1993, 2000) and Cramer (2002) studied this event in a borehole from Clayton, NJ, providing clay mineralogic and isotopic data (Fig. F10). Drilling at Bass River and Ancora preserves an exceptional record of the PETM with a chronology provided by integrated nannofossil, planktonic foraminiferal, and magnetostratigraphy; these neritic sections rival the best deep-sea sections in providing a complete record of these global climatic events (Cramer et al., 1999, 2000; Cramer, 2002). Isotopic analyses at all three boreholes show a rapid (<10 k.y.) negative shift in 13C and 18O values (~4 in carbon and ~2 in oxygen), and clay mineralogical analyses for Bass River and Clayton show a sharp increase in kaolinite coincident with the isotopic shift (Fig. F10) (Cramer et al., 1999; Cramer, 2002). Stable isotopic values remain low and kaolinite content remains high throughout a thick section above the CIE in these neritic sections. Cramer et al. (1999) interpreted these data as indicative of an abrupt shift at the time of the CIE to a warmer and wetter climate that persisted for >300-400 k.y. along the North American Mid-Atlantic coast, unlike deep-sea records that show an exponential return to pre-CIE conditions after ~200 k.y. (Katz et al., 1999, Rohl et al., 2000). The thick interval of low isotopic values at Bass River, Ancora, and Clayton coincides with an interval magnetically dominated by <100-nm-diameter magnetite grains. The magnetically anomalous material has been interpreted as derived from a compact impact plume condensate, leading to speculation that the P/E boundary event may have been triggered by a major impact event and reinterpretation of the thickness of this interval on the NJ coastal plain as due to rapid redeposition of impact ejecta over <20 k.y. rather than persistent warm conditions following the PETM (Kent et al., 2001; Cramer, 2002; Kent et al., unpubl. data).
Earliest Oligocene Event
Kominz and Pekar (2001) provided a backstripped estimate of 54 ± 10 m of eustatic lowering that can be correlated (Pekar et al., 2001) with the well-known earliest Oligocene global 18O increase (e.g., Miller et al., 1987). This implies development of an ice sheet that was as large, if not slightly larger, than the modern East Antarctica ice sheet, consistent with the results of Miller et al. (1987) and Zachos et al. (1994). Nevertheless, the large size of the earliest Oligocene (~33.8 Ma) ice sheet is surprising, especially considering its near disappearance about a million years later: Kominz and Pekar's (2001) eustatic estimate shows a 46 ± 15-m eustatic rise at 32.7 Ma, and records from Antarctica show the return of alpine Nothofagus trees at about this time to the Ross Sea, Antarctica (Cantrill, 2001). Transient ice sheets returned numerous times during the Oligocene-middle Miocene (e.g., Barrett, 1999), and eustatic estimates show numerous large rises and falls (Pekar et al., 2001; Miller et al., 1998a), testifying to a very dynamic ice sheet between ~33 and 14 Ma. By the middle Miocene, the East Antarctic ice sheet had become a permanent feature (e.g., Barrett, 1999) and ice volume changes were primarily controlled by small changes in Antarctica and the nascent growth of Northern Hemisphere ice sheets (Wright, 1998).
Drilling as part of Leg 174AX (Figs. F11, F12, F13, F14) was very successful in addressing local water resource issues. The Bass River and Ancora boreholes targeted Cretaceous aquifers in the Mount Laurel, Englishtown, and Potomac-Raritan-Magothy (PRM) Formations (see Zapecza, 1989, for discussion of these aquifers). Ocean View targeted Miocene aquifers (e.g., the Atlantic City 800-ft sand aquifer of Zapecza, 1989) and confining units between Cape May and Atlantic City. Bethany Beach targeted the hydrogeology of the locally important Pocomoke, Ocean City, and Manokin aquifers, especially in delineating the distribution of freshwater and saline-water zones deeper in the subsurface (see Andres, 1986, for discussion of these aquifers). Continuous coring has shown that aquifer-confining unit couplets are sequences, bounded by unconformities (e.g., Sugarman and Miller, 1997). The upper HST sands comprise important aquifers in both regions that are generally confined by the overlying TST or lower HST. Thus, sequence stratigraphy provides a means to predict the continuity and regional distribution of aquifer-confining bed units (Sugarman and Miller, 1997). Coring has helped resolve issues with the updip-downdip and along-strike relationships of aquifer-confining bed units. For example, the Atlantic City 800-ft sand near Atlantic City is comprised of two sand bodies that make up the HSTs of the Kw1a and Kw1b sequences (Sugarman and Miller, 1997). However, at Cape May, there are two to three sand bodies that could be mapped as the Atlantic City 800-ft sand, with the highest of these associated with the Kw1c sequence (Miller et al., 1996a). Drilling at Ocean View shows that the Kw1c sequence pinches out between Cape May and Ocean View and thus the upper sand at Cape May is not equivalent to the upper sands at Ocean View, Atlantic City, and points north. The drilling also makes clear the control of depositional facies on aquifer architecture. In DE, aquifer sand units developed in HST shoreface deposits are areally extensive and can be correlated tens of kilometers from the Bethany Beach borehole. These include the middle and lower Miocene aquifer units encountered at Bethany Beach: the Milford aquifer, which appears to represent the upper HST of Kw2a based on current interpretation of the Sr age control; and the Cheswold aquifer, which is interpreted as the upper HST of the Kw1a. Both can be correlated on geophysical logs through most of southern and central DE. In contrast, aquifer units developed in estuarine-barrier facies, such as the upper? Miocene Pocomoke and Ocean City aquifers at Bethany Beach, are laterally discontinuous, reflecting the high degree of facies change in these environments. The amount of sand present in these estuarine-barrier intervals varies even on the kilometer scale, making detailed local correlation difficult.
Pore water geochemical studies of onshore sites provide important constraints on the role of confining units (see Leg 150X studies by Szabo et al., 1997; Pucci et al., 1997). Pore waters of the lower Miocene confining units at Bass River were geochemically typed and compared with adjacent aquifers (Reilly, 2001). Pore water in the confining units are geochemically distinct from adjacent aquifers and correlative confining units at Atlantic City, 15 km to the south.
