BIOSTRATIGRAPHIC DISCUSSION
Biocalcareous Stratigraphic Events
The biostratigraphy afforded by calcareous nannofossils in material from Leg 178 continental rise sites is discontinuous and incomplete. Nevertheless, they allow biostratigraphic age control when there would otherwise be only paleomagnetic data, as diatoms, radiolarians, and foraminifers are rare and reworked in the upper intervals of these sites. Overall abundance and diversity is often low, with several samples having abundant or common placoliths (Tables T2, T3). Table T4 lists the biostratigraphic events observed during this study.
The acme of E. huxleyi in the uppermost Holocene is not observed in any of the samples examined from these three holes (1096A, 1096B, and 1101A). The base of E. huxleyi is noted in Hole 1101A (interval 178-1101A-2H-5, 90 cm, to 2H-CC [15.6-18.2 mbsf]), allowing the CN15/NN21a biozone boundary to be assigned in this hole. In all three sites the next 10-15 m of sediment below this interval is barren of any nannofossils. Holes 1101A and 1096B have samples with only scarce or rare occurrences down through 72 and 78 mbsf, respectively. Hole 1096A contained two samples with higher abundances at 28 and 53 mbsf, the first of which contained the youngest sample, with common to abundant numbers of Coccolithus pelagicus. The samples containing greater numbers (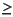 1 per field of view) are indicated with a dot on Figures F2 and F3. The gray bars on these two figures illustrate the periodic nature of the samples containing calcareous nannofossils. The overall abundance of nannofossils is also indicated with the graph on the right side of the figures.
1 per field of view) are indicated with a dot on Figures F2 and F3. The gray bars on these two figures illustrate the periodic nature of the samples containing calcareous nannofossils. The overall abundance of nannofossils is also indicated with the graph on the right side of the figures.
The next biostratigraphic event observed is the LO of Pseudoemiliania lacunosa (interval 178-1096A-3H-CC to 4H-2, 40 cm [25.23-28.6 mbsf]). The large barren interval in Hole 1101A makes it unwise to place this event in this hole. The LO of P. lacunosa indicates the base of Biozones CN14b/NN20. Hole 1096B contains the next two stratigraphically older events, the LO of Reticulofenestra asanoi and the reentrant of medium-sized Gephyrocapsa spp. These two events are contained in one sample at 78.41 mbsf (Section 178-1096B-9H-6, 61 cm), with the ranges for each extending to the samples on either side (up- and downcore). The LO of R. asanoi ranges between 76.76 and 78.41 mbsf (interval 178-1096B-9H-5, 46 cm, to 9H-6, 61 cm) and the reentrant of medium-sized Gephyrocapsa spp. between 78.41 and 79.72 mbsf (interval 178-1096B-9H-6, 61 cm, to 9H-7, 42-44 cm). Raffi et al. (1993) observed the reentrant event to occur at 0.96 Ma. Both of these events occur between 1.03 and 1.05 Ma in Hole 1096B (based on paleomagnetics and sedimentary rates at this site). The difference in the event(s) age and the fact that there are barren and low-abundance samples upcore from Sample 178-1096B-9H-6, 61 cm, indicate that neither of these are likely to be the actual LO and reentrant events. They are both included in the table of calcareous nannofossil events despite this possibility, due to the uncertainty of how calcareous nannofossil events occurred in the Southern Ocean and the fact that Hole 1101A also contained the reentrant event of medium-sized Gephyrocapsa spp. between 1.05 and 1.15 Ma (Table T4).
The FO of R. asanoi was noted at both sites, occurring at 90.4-91.5 mbsf (interval 178-1096B-11H-1, 11 cm, to 11H-2, 70 cm) and 84.77-84.94 mbsf (interval 178-1101A 10H-6, 107 cm, to 10H-CC). The total range of the large variety of Gephyrocapsa spp. was observed at both sites: from 91.5-97.42 to 128.84-131.49 mbsf at Site 1096 and from 84.94-90.9 to 94.19-98.88 mbsf at Site 1101 (see Tables T2 and T3 for samples). Neither Gephyrocapsa oceanica or Gephyrocapsa caribbeanica (large varieties) occurred in great numbers in either hole, yet these species were distinctive in both their appearance and in the intervals in which they did occur, with very little reworking indicated.
The LO of Helicosphaera selli was observed only at Site 1101 at 90.9-93.87 mbsf (interval 178-1101A-11H-4, 70 cm, to 11H-6, 67 cm). The base of Biozone CN13b is defined by the FO of medium-sized Gephyrocapsa spp., noted in Site 1096 at 138.37-138.45 mbsf (interval 178-1096B-15H-5, 11 cm, to 15H-CC). Calcareous nannofossils were not observed (downhole) at 168.37 and 121.1 mbsf in Holes 1096B and 1101A, respectively.
As can be seen in Figures F2 and F3, the total abundance and frequency of occurrence of calcareous nannofossils increase toward the lower part of their range at each site. Many of the samples with high total abundance contain large numbers of C. pelagicus (these samples are illustrated by black dots in the aforementioned figures). It is worthy of note that in Hole 1096B, maximum abundances of C. pelagicus are associated with lower numbers of medium-sized Gephyrocapsa spp. Sato et al. (1998) studied C. pelagicus from middle and high northern latitudes in relation to its significance as a cold-water current indicator. Presently, this species occurs only in high-latitude regions but has a fossil distribution as far south as the equatorial Pacific (>4.5, 4.5-3.66, and 2.75-1.65 Ma). This southerly distribution was associated with the elevation of the Isthmus of Panama, causing the cold northern currents to flow into the equatorial Pacific. Wei and Wise (1992b) noted C. pelagicus to mark the warmer end of the spectrum when compared with an even colder water species, Reticulofenestra perplexa, from the Antarctic during the Miocene.
Pliocene and Pleistocene occurrences of calcareous nannofossils in ODP cores were noted previously from the Southern Ocean during Legs 113, 119, and 120 (Barker and Kennett, 1988; Barron, Larson, et al., 1989; Barron et al., 1991; Wei and Wise, 1990, 1992a, 1992b). Hole 693A (Leg 113) contains rare occurrences of G. oceanica in Section 113-693A-1H-CC, which were believed at the time to be a contaminant (Wei and Wise, 1990). This site is situated in a hemipelagic setting in the eastern Weddell Sea. Site 690 (also Leg 113) contained rare occurrences of C. pelagicus and P. lacunosa. Wei and Wise (1990) also concluded that the presence of C. pelagicus at Site 690, in the middle Pliocene to upper Pleistocene, suggests that interglacial conditions during this time were sufficiently warm to allow this species to exist at this site. Hole 747A (Leg 120) contained abundant to common E. huxleyi in the upper two samples. Several samples within the Pliocene-Pleistocene interval had rare to abundant occurrences of small Gephyrocapsa spp., with one sample (120-747A-1H-4, 10-11 cm) containing abundant G. oceanica. C. pelagicus is rare to very abundant throughout the Pliocene-Pleistocene interval of this Hole, with a somewhat discontinuous range (Wei and Wise, 1992a). Wei and Thierstein (1991) (Leg 119) observed an early to middle Pleistocene assemblage in Sample 119-738A-1H-CC (C. leptoporus, C. pelagicus, G. caribbeanica, Helicosphaera kamptneri, and P. lacunosa). They suggested that the presence of these species might indicate surface waters that were significantly warmer during this period than they are today.
Villa and Wise (1998) observed Thoracosphaera spp. in the CRP-1 hole of the Cape Roberts Project in the Ross Sea. This is interpreted to mean several possible things. Either the ability of this genus to form cysts allowed them to survive in the Antarctic environment, or there were warmer conditions in the Ross Sea during the time of production and deposition (or a bit of both). In addition to this core, they also examined others from various sites around Antarctica. Several Deep Freeze cores, also from the Ross Sea, were examined and found to contain rare occurrences of C. pelagicus, Reticulofenestra producta, Gephyrocapsa spp., and Thoracosphaera spp. A SEDANO (Sediment Drifts of the Antarctic Offshore) core nearer to the area of Leg 178, the Pacific margin of the Antarctic Peninsula, contained Gephyrocapsa ericsonii, C. leptoporus, C. pelagicus, Helicosphaera spp., and E. huxleyi.
Burckle and Pokras (1991) report finding early Holocene calcareous nannofossils in cores near the western side of the Antarctic Peninsula. In this same paper, however, they conclude that average surface-water temperatures have to be above 3°C during the growing season to produce and preserve calcareous nannofossils. Following this approach, Kennett and Hodell (1993, 1995) argue that the absence of Pliocene calcareous nannofossils in Antarctic waters can be used as a paleotemperature index; the absence of calcareous nannofossils must indicate temperatures below 5°C. Under such warming, they surmise, biosiliceous sedimentation in Antarctic waters would be replaced by biocalcareous sedimentation, such as that seen north of the Polar Front today. An obvious problem with generalizations is that in some instances they do not completely apply. At this time, there is no easy explanation why calcareous nannofossils were living and subsequently being deposited in the hemipelagic drifts on the western Pacific margin of the Antarctic Peninsula during the Pleistocene. There were no concrete conclusions reached in previous investigations that observed Pleistocene and Pliocene nannofossils in the studies mentioned earlier from several points in the Southern Ocean. That they were either not growing or not being deposited in large numbers is quite evident. The most common species observed is not always the most dissolution resistant (i.e., C. pelagicus), yet there are intervals where this species is abundant. Many of the samples are poorly preserved, but not completely.
These biocalcareous components of the sediment in close proximity to the continent can not and should not be ignored or considered to be random mistakes; they occur in too high a frequency and diversity to be such.
