MICROBIOLOGY
Microbiological sampling of Site 1229 covered the sulfate-reducing zone near the sediment/water interface, the sulfate-depleted methanogenic zone between ~35 and 90 mbsf, and the sulfate-rich deep layer at depths between 90 and 186 mbsf, which was influenced by the presence of a deep subsurface brine. Whenever possible, two sections of each core from the upper 100 m of the sediment column were routinely sampled for deoxyribonucleic acid (DNA) analysis, measurements of sulfate reduction rates, hydrogen concentration and turnover, methanogenesis rates, acetate turnover, thymidine incorporation, bacterial lipid biomarkers, adenosine triphosphate, fluorescent in situ hybridization (FISH), and iron/manganese/sulfur solid-phase geochemistry (Fig. F6) (Cores 201-1229A-2H through 11H). Poor recovery of Core 201-1229A-7H (49.5-59 mbsf) led to this depth interval being substituted with two core sections from the same depth interval of Hole 1229D (Sections 201-1229D-7H-1 and 7H-4) (Fig. F7). Every core that could be obtained from the deeper layers of the sediment column was sampled to analyze the prokaryotic communities and activities in the deep sulfate-rich layer (Sections 201-1229A-12H-3 through 22H-2) (Fig. F6).
Sampling in Hole 1229D focused on fine-scale resolution of intervals that had been missed or inadequately sampled in Hole 1229A. Sampling in five sections of the mudline core in Hole 1229D (Sections 201-1229D-1H-1,1H- 2, 1H-3, 1H-4, and 1H-5) aimed at fine resolution of the highly compressed chemical gradients at the top of the sediment column in order to provide good end-member data for the microbiological and geochemical analyses in deep sediment layers (Fig. F7). Sampling in Sections 201-1229D-4H-3, 4H-4, and 4H-5 aimed at resolving the low-sulfate interval between 30 and 33 mbsf, where sulfate declines to <1 mM (Figs. F6, F7). The two bottom sections of the short Core 201-1229D-12H (Sections 12H-2 and 12H-3) and the consecutive two top sections of Core 13H (Sections 13H-1 and 13H-2) were sampled to obtain a fine resolution in the lower sulfate-methane transition zone between 85 and 90 mbsf (Fig. F5) that had not been sampled sufficiently in Cores 201-1229A-10H and 11H (Fig. F7). In addition, specific samples were recovered in Hole 1229D from defined depths that had not been sampled in sufficient detail at similar depths in Hole 1229A (Sections 201-1229D-2H-5, 6H-1, 9H-4, and 15H-3).
Total Prokaryotic Cell Enumeration
During core processing in the 10°C reefer, sample plugs (1 cm3) for total prokaryotic cell enumeration were taken from a total of 24 depths between 8.88 and 185.7 mbsf in Hole 1229A (18 samples) and between 0.01 and 53.86 mbsf in Hole 1229D (6 samples). Prokaryotes were present in all samples to 185.7 mbsf (Fig. F8). The highest number was found at 90.45 mbsf (Sample 201-1229A-11H-2, 5-11 cm), which contained 9.5 x 109 cells/cm3. The lowest number was at the base of the hole, with 2.7 x 106 cells/cm3, 3500-fold lower than at 90.45 mbsf.
The overall depth profile of cell numbers per cubic centimeter follows a trend observed at other ODP sites (Parkes et al., 1994), with many of the data lying very close to, or on, the mean regression line (Fig. F9). Two zones of major interest, just above the 35-mbsf upper sulfate-methane transition and the lower sulfate-methane transition at ~90 mbsf (see "Biogeochemistry") were intensively sampled. Prokaryotic cell numbers were very high at these depths and in both cases exceeded the upper 2-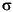 envelope of the existing database. The upper sulfate-methane transition zone showed elevated counts between 20.98 mbsf (Sample 201-1229A-3H-5, 58-60 cm) and 40.35 mbsf (Sample 5H-5, 95-100 cm), with a maximum of 1.18 x 108 cells/cm3 at 29.81 mbsf (Sample 201-1229D-4H-3, 101-107 cm). This represented a tenfold increase over predicted cell counts at that depth. The increase in cell numbers at the lower sulfate-methane transition zone was broader, lying between 84.48 mbsf (Sample 201-1229A-10H-4, 58-64 cm) and 157.98 mbsf (Sample 18H-2, 108-114 cm). The lower boundary may represent a sampling artifact, since data points between 125 and 158 mbsf were not available. Within this broad band of increased cell numbers, a peak was present at 90.45 mbsf (Sample 201-1229A-11H-2, 5-11 cm) with 9.48 x 109 cells/cm3, which represented a 1730-fold increase over predicted cell counts at that depth. This value was exceptionally high for a deep marine sediment. To our knowledge, this is the highest prokaryotic cell count ever found well below the seafloor. To exclude potential sources of error and bias, the counts were repeated. Solutions were refiltered, new sterile equipment was used, blanks were checked, and a different person redid the count. The original enumeration was confirmed.
envelope of the existing database. The upper sulfate-methane transition zone showed elevated counts between 20.98 mbsf (Sample 201-1229A-3H-5, 58-60 cm) and 40.35 mbsf (Sample 5H-5, 95-100 cm), with a maximum of 1.18 x 108 cells/cm3 at 29.81 mbsf (Sample 201-1229D-4H-3, 101-107 cm). This represented a tenfold increase over predicted cell counts at that depth. The increase in cell numbers at the lower sulfate-methane transition zone was broader, lying between 84.48 mbsf (Sample 201-1229A-10H-4, 58-64 cm) and 157.98 mbsf (Sample 18H-2, 108-114 cm). The lower boundary may represent a sampling artifact, since data points between 125 and 158 mbsf were not available. Within this broad band of increased cell numbers, a peak was present at 90.45 mbsf (Sample 201-1229A-11H-2, 5-11 cm) with 9.48 x 109 cells/cm3, which represented a 1730-fold increase over predicted cell counts at that depth. This value was exceptionally high for a deep marine sediment. To our knowledge, this is the highest prokaryotic cell count ever found well below the seafloor. To exclude potential sources of error and bias, the counts were repeated. Solutions were refiltered, new sterile equipment was used, blanks were checked, and a different person redid the count. The original enumeration was confirmed.
The peak in prokaryotic numbers at 90.45 mbsf was present within a prominent gray homogeneous quartz- and feldspar-rich clay (see "Subunit
IB" in "Description of Lithostratigraphic Units" in "Lithostratigraphy"). To confirm the high prokaryotic cell counts, additional subsamples were taken from two other independently sampled sources within the same clay zone for enumeration. These were a 0.2-mL slurry sample from 91.14 mbsf (Sample 201-1229A-11H-2, 88-93 cm) and a 1-cm3 whole-round core (WRC) sample from 91.28 mbsf (Sample 11H-2, 103-109 cm). These gave prokaryotic cell counts of 5.81 x 108 and 2.06 x 109 cells/cm3, respectively (Figs. F8, F9). The mean prokaryotic cell count density in this narrow 83-cm band amounts to 2.25 x 109 cells/cm3, a 410-fold increase over the mean cell number from all previously published sites at this depth.
Numbers of dividing cells (suggested as an index of growth activity) are typically <10% of the total count. As expected, dividing cells, as a percentage of the total count, are high near the surface (Fig. F8). They decrease to a minimum at 20.98 mbsf (Sample 201-1229A-3H-5, 58-60 cm), before increasing again. Interestingly, the maximum percentage of dividing cells around the upper sulfate-methane transition zone is present at 40.35 mbsf (Sample 201-1229A-5H-5, 95-100 cm), well below the maximum direct count. In all cases where a reliable quantification was possible, the proportion of dividing cells amounted to ~10% of the total counts below the upper sulfate-methane transition zone and also at the prokaryotic cell abundance peak at the deeper sulfate-methane transition zone.
The high amount of organic matter and the silty clay with very small particle size made direct counting at this site more problematic than usual. Generally, subsamples of 15-40 µL can be processed, but in this case subsample size was restricted to a maximum of 10-12 µL. Consequently, the detection limit increased to 5.8 x 105 cells/cm3. The percentage of dividing cells should be viewed with caution where the direct count is <1 x 107 cells/cm3.
While drilling cores for microbiology, the potential for contamination with bacteria from the surface is highly critical. Contamination tests were continuously conducted using solutes (PFT) or bacterial-sized particles (fluorescent microspheres) to check for the potential intrusion of drill water from the periphery toward the center of cores and thus to confirm the suitability of the core material for microbiological research. We used the chemical and particle tracer techniques described in ODP Technical Note 28 (Smith et al., 2000). Furthermore, the freshly collected cores were visually examined for possible cracks and other signs of disturbance by observation through the transparent core liner. Core sections observed to be disturbed before or after subsampling were not analyzed further. Such disturbance phenomena are critical to the integrity of the core material and therefore also to its usefulness for microbiological studies.
Perfluorocarbon Tracer
PFT was injected continuously into the drilling fluid during drilling of Holes 1229A and 1229D (see "Perfluorocarbon Tracer Contamination Tests" in "Microbiological Procedures and Protocols" in "Microbiology" in the "Explanatory Notes" chapter). PFT concentration analyses focused on microbiology cores and especially on sections that were used for slurry preparation and cultivations. To compare the PFT concentrations in the center of a core to the PFT concentrations at the periphery of the same core, a 5-cm3 subcore sample was also taken at the core periphery, adjacent to the core liner. Whenever possible, the samples were taken directly on the catwalk because the PFT content of catwalk air was usually not detectable.
Low levels of potential seawater contamination (Table T5) were found for the center portions of all tested cores of Holes 1229A and 1229D. With one exception (Section 201-1229A-6H-2), the values were not higher than 0.07 µL seawater/g sediment (average = 0.039 µL seawater/g sediment). The outer portions of all tested cores had a significantly higher level of PFT tracer and potential seawater contamination. In all cases, the PFT content and the estimated potential seawater contamination levels were higher in the periphery of the core than in the center (see House et al., this volume).
Of the eight master slurry samples taken from Site 1229 (Table T6), only the slurries from Cores 201-1229A-9H and 11H showed small concentrations of PFT (~0.06 µL potential seawater contamination/mL slurry).
Assuming 5 x 108 prokaryotic cells/L surface seawater, each 0.1 µL seawater contamination may represent as many as 50 contaminating cells if the sediment is porous enough to allow cells to travel with the PFT.
Particulate Tracer
Fluorescent microspheres (beads) were deployed on all eight cores from which slurries were made at this site. For each slurry, two subsamples were processed: (1) a sample of the slurry to check contamination and (2) a scraping from the outer surface of the core to confirm deployment of microspheres.
Microsphere deployment was confirmed from the outer core scrapings in all six cores that were sampled for slurry preparation (Table T7). Microspheres were detected in two slurry samples. In Sample 201-1229A-3H-2, 40-54 cm, a single bead was observed; further microscopic searching of the membrane failed to detect any additional beads. It is believed that a single microsphere represents filter handling and processing effects rather than contamination (see "Fluorescent Microparticle Tracer" in "Procedures and Protocols" in "Microbiology" in the "Explanatory Notes" chapter). In Sample 201-1229A-11H-2, 74-88 cm, however, three microspheres were observed within the counting grid and many more microspheres were present outside of the counting area. It therefore seems likely that this sample was contaminated.
Since the complex geochemical profiles of Site 1229 (Fig. F5) indicate distinct zones of contrasting microbial activities, slurry samples from nine different depths were used for cultivation experiments (Table T8). These slurries included the eight master slurries checked for contamination (Table T7) and an additional slurry from Core 201-1229D-15H for enrichments of predominantly thermophiles. Slurries for cultivation were prepared by subcoring with two 60-mL syringes from the center of two freshly broken surfaces after precutting the core liner with the ODP cutter. This technique provided untouched (although not always smooth) surfaces that were immediately sampled. The inoculated media were selective for fermentative prokaryotes, sulfate reducers, methanogens, and various anaerobic chemolithoautotrophic and heterotrophic prokaryotes that use iron(III) or manganese(IV) as an electron acceptor. Quantitative cultivations (most probable number [MPN] experiments) and enrichment cultures were started at temperatures between 15° and 80°C. For this site, additional microbiological work was undertaken considering the following aspects:
- To quantify culturable methanogens at different depths in the sediment column, dilution series were prepared using substrates that are characteristic for major physiological and phylogenetic groups of methanogens. Two of these substrates, methyl amines (typically found in nearshore environments) and hydrogen/carbon dioxide can also be used by acetogens. Potentially, acetogens could provide acetate for the sulfate reducers at the sulfate/methane interface.
- In order to study the role of sulfate as a limiting electron acceptor, both sulfate-free and sulfate-containing media were used for inoculation of samples from the sulfate-containing sediment layers, the sulfate-methane transition zone, and sulfate-free sediment layers. Whereas the normal sulfate-containing media were reduced with FeS, the sulfate-free media were reduced with titanium citrate and were kept sulfide-free to exclude sulfide oxidation to sulfate.
- A considerable proportion of subsurface prokaryotes could be spore-forming bacteria. Spores are bacterial resting stages that are resistant to environmental stress factors, such as nutrient depletion, desiccation, heat, and irradiation. To quantify the contribution of spore-forming bacteria to the total cultivable prokaryotic community, pasteurized (heated to 80°C for 10 min) and untreated slurries were used in parallel for MPN experiments. All shipboard cultivation experiments and the media and incubation temperatures are shown in Table T8. For details of the various media compositions and incubation conditions see Tables T4, T5, and T7, and "Procedures and Protocols" in "Microbiology" in the "Explanatory Notes" chapter.
13C substrate incubations were initiated for postcruise analysis by fluorescence in situ hybridization-secondary ion mass spectrometry (FISH-SIMS) using material from Cores 201-1229A-2H, 3H, 4H, 6H, and 11H. In each case, 10 mL of the master slurry was injected into each bottle. The 13C substrates used were methane, acetate, and glucose. For Cores 201-1229A-4H and 11H, an additional acetate bottle was inoculated, and for Core 11H, no glucose bottle was used.
