MICROBIOLOGY
Microbiological samples were taken from the second and fifth section of every available core between Cores 201-1227A-2H and 6H. This high-resolution microbiological sampling covered the sulfate-reducing zone and extended across the sulfate/methane interface into the methanogenic zone (see "Biogeochemistry"). Beginning with Core 201-1227A-7H, every undisturbed core was sampled at a frequency of one section per core. In Hole 1227D, additional samples were obtained for microbiological and biogeochemical study from selected depths (Figs. F8, F9). The uppermost five sections of Core 201-1227D-1H were sampled to increase resolution near the sediment/water interface. Cores 201-1227D-4H, 5H, and 8H were sampled to fill any gaps remaining from our sample distribution from Hole 1227A. Higher-resolution sampling of Cores 201-1227D-4H and 5H focused on the sulfate/methane transition between 35 and 43 mbsf. Core 201-1227A-14H had a highly porous and spongy structure, presumably a result of gas-induced core expansion.
The sampling program for each section is shown in detail for Hole 1227A in Figure F8 and for Hole 1227D in Figure F9. For deoxyribonucleic acid (DNA), fluorescence in situ hybridization (FISH), adenosine triphosphate (ATP), lipid biomarker analysis, solid-phase iron/manganese/sulfur chemistry, and solid-phase phosphate chemistry, samples were fixed or frozen, as appropriate, for processing on shore (for details see "Procedures and Protocols"). Whole-round cores (WRC) were subsampled into 5-mL syringes for subsequent measurements of hydrogen concentrations and for rates of tritium turnover, sulfate reduction, methanogenesis, acetate turnover, and thymidine incorporation into DNA. Samples for acridine orange direct counts were fixed from the same sections, and the cells were counted on the ship. Samples for anaerobic oxidation of methane experiments, sulfate reduction experiments, 18O isotope analysis, pyrolysis of organic material, and diffusion experiments were taken at several defined intervals (Fig. F8). Samples at ostensibly dolomite-bearing layers (Sections 201-1227A-3H-5, 8H-2, and 12H-2; confirming analysis will be done postcruise) were taken for FISH, enrichment, lipid biomarker, and sedimentological analyses. From Section 201-1227D-5H-5, where a barium peak was observed in the interstitial water profile from Hole 1227A (see "Biogeochemistry"), extra samples were obtained for shore-based sulfate reduction experimentation (using radiolabeled Ba35SO4), DNA analysis, and shore-based microscopic studies and enrichment cultures on barite.
Samples for cultivation were taken from the near-surface sediment column (Sections 201-1227D-1H-1 and 201-1227A-2H-5 and 3H-5), from intermediate depths near the sulfate-methane transition (Sections 201-1227A-5H-5, 201-1227D-4H-6, and 201-1227A-7H-2), and from the deeper methanogenic portion of Hole 1227A (Sections 12H-2 and 18H-2). In order to study the role of sulfate as a biologically limiting factor, sulfate-free and sulfate-containing media were used in samples from intervals with and without sulfate.
Total Bacterial Enumeration
Samples of 1-cm3 plugs for total prokaryotic enumeration were taken on the catwalk from a total of 21 depths between 8.4 and 144.3 mbsf in Hole 1227A (15 samples) and between 0.15 and 38.8 mbsf in Hole 1227D (6 samples). Additionally, 2-mL samples of 25% slurry (vol/vol) were taken from six slurries prepared in the laboratory (see "Preparation of Slurries" in "Procedures and Protocols" in "Microbiology" in the "Explanatory Notes" chapter). Prokaryotic cells were present in all enumerated samples taken from the catwalk to 144.3 mbsf (Fig. F10). The highest number of prokaryotes was found in the near-surface sample (Sample 201-1227D-1H-1, 15-16 cm), which contained 1.42 x 107 cells/cm3. The lowest number of prokaryotes was at the base of the hole, with 1.77 x 106 cells/cm3, an eightfold decrease.
The overall depth profile of cell numbers per cubic centimeter follows a trend observed at other ODP sites (Parkes et al., 1994). However, cell numbers are persistently lower than the average from previous sites (Fig. F11). All data except the shallowest sample depth fit within the 2-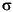 envelope around the fit of total censused data from previous ODP sites. Counting has so far focused on the sulfate-methane transition zone, between ~35 and 43 mbsf (see "Biogeochemistry"). Cell numbers were maximal at 31.3 mbsf (Sample 201-1227A-4H-5, 105-110 cm), with 1 x 107 cells/cm3 (Fig. F10). Two-sample t-tests established that the numbers of prokaryotes at this depth were significantly greater than those from the sample directly above (t = 3.91; n = 6; P < 0.05) at 25.95 mbsf (Sample 201-1227A-4H-1, 135-140 cm) and from the sample directly below (t = 5.18; n = 6; P < 0.01) at 38.4 mbsf (Sample 5H-3, 130-135 cm).
envelope around the fit of total censused data from previous ODP sites. Counting has so far focused on the sulfate-methane transition zone, between ~35 and 43 mbsf (see "Biogeochemistry"). Cell numbers were maximal at 31.3 mbsf (Sample 201-1227A-4H-5, 105-110 cm), with 1 x 107 cells/cm3 (Fig. F10). Two-sample t-tests established that the numbers of prokaryotes at this depth were significantly greater than those from the sample directly above (t = 3.91; n = 6; P < 0.05) at 25.95 mbsf (Sample 201-1227A-4H-1, 135-140 cm) and from the sample directly below (t = 5.18; n = 6; P < 0.01) at 38.4 mbsf (Sample 5H-3, 130-135 cm).
Numbers of dividing cells (suggested as an index of growth activity) are typically <10% of the total count. As expected, dividing cells, as a percentage of the total count, are high near the surface (Fig. F10) and then decrease steadily to a minimum at 38.4 mbsf (Sample 201-1227A-5H-3, 130-135 cm). The two data points below this depth suggest that dividing cells increase at greater depth; however, there are insufficient data here to justify any conclusions. The high amount of organic matter in the sediment at this site made direct counting more problematic than usual. Where subsamples of 15-40 µL can generally be processed, in this case, subsample size was restricted to a maximum of 12 µL. Consequently, the detection limit increased to 1.3 x 105 cells/cm3 (Fig. F10) and thus counting sensitivity decreased.
While drilling cores for microbiology, the potential for contamination with bacteria from the surface is highly critical. Contamination tests were continuously conducted using solutes (perfluorocarbon tracer [PFT]) or bacterial-sized particles (fluorescent microspheres) to check for the potential intrusion of drill water from the periphery toward the center of cores and thus to confirm the suitability of the core material for microbiological research. We used the chemical and particle tracer techniques described in ODP Technical Note 28 (Smith et al., 2000). Furthermore, the freshly collected cores were visually examined for possible cracks and other signs of disturbance by observation through the transparent core liner. Core sections observed to be disturbed before or after subsampling were not analyzed further. Such disturbance phenomena are critical to the integrity of the core material and therefore also to its usefulness for microbiological studies.
Perfluorocarbon Tracer
The perfluorocarbon tracer (PFT) was injected continuously into the drilling fluid during drilling of Holes 1227A and 1227D (see "Perfluorocarbon Tracer Contamination Tests" in "Procedures and Protocols" in "Microbiology" in the "Explanatory Notes" chapter). PFT sampling focused on microbiology cores and especially on sections that were used for slurry preparation and cultivations. To increase the sensitivity of PFT detection, two (instead of one) 5-cm3 subcores were taken from the center of a section and pooled for PFT measurement at Site 1227. This strategy was used at all further sites of Leg 201. The increased amount of sample material resulted in an improved detection of PFT (corresponding to ~0.01 µL seawater contamination/g sediment), under the reasonable assumption that the PFT signal follows proportionally the twofold increased sample volume. In parallel, samples were taken from the outer portion of each sediment core next to the core liner to assess the difference in contamination potential between inner and outer portions of a core. Whenever possible, the samples were taken directly on the catwalk because the PFT content of catwalk air was usually not detectable.
Low levels of potential seawater contamination (Table T5) were found for the center portions of all tested cores of Holes 1227A and 1227D and did not exceed 0.05 µL seawater/g sediment (average = 0.021 µL seawater/g sediment). The outer portions of all tested cores had a significantly higher level of PFT tracer and potential seawater contamination (average = 1.77 µL seawater/g sediment). In all cases, the PFT content and the estimated potential seawater contamination levels were higher at the periphery of the core than in the center. Two samples showed seawater contamination levels of more than double the average from Sections 201-1227A-7H-2 and 201-1227D-4H-6, indicating high variability and patchiness for the more highly contaminated areas in the outer core.
Of the six master slurry samples taken from Site 1227 (Table T6), only the slurry from Core 201-1227A-3H showed small concentrations of PFT (~0.03 µL potential seawater contamination/g slurry). This value is near the detection limit of 0.025 µL potential seawater contamination/mL slurry. Assuming 5 x 108 prokaryotic cells/L surface seawater, each 0.1 µL seawater contamination may represent as many as 50 contaminating cells if the sediment is porous enough to allow cells to travel with the PFT.
Particulate Tracer
Fluorescent beads were deployed on seven occasions at this site, including all cores from which slurries were made. On two occasions the bead bag failed to burst. The first time this happened (Core 201-1227A-5H), it remained unrecognized until well after slurry processing had finished. At the second occurrence, the failure was recognized immediately and beads were run on the following core, and the slurry was made on the latter core (Section 201-1227A-12H-2). Modifications to the delivery mechanism for bead bags were made at the next site. For each slurry, the presence of beads was checked in up to three subsamples: (1) a sample of the slurry; (2) when necessary, a sediment sample tenfold diluted in 2% formaldehyde (used also for direct prokaryotic counts); and (3) a scraping from the outer surface of the core (processed to confirm deployment of beads). Beads were not detected in any of the six slurries. The presence of beads was confirmed from the outer core scrapings in all slurries (not in Core 201-1227A-5H), confirming deployment. Consistent with the low PFT values, no beads were detected in any of the slurries from Site 1227.
Vertical profiling of geochemical parameters indicated that the sulfate-methane transition is present from 35 to 43 mbsf (see Fig. F6). Accordingly, the presence of prokaryotes possibly responsible for sulfate reduction, acetogenesis, and methanogenesis throughout the sulfate-methane transition were evaluated by a combined approach of enrichment cultures and most probable number assays. Samples for slurries were taken from five different depths in Hole 1227A, including the sulfate-methane transition zone, and from two samples in Hole 1227D (Table T7). The slurry samples were taken by subcoring with two 60-mL syringes from the center of two freshly broken sediment core surfaces. The slurry was used to inoculate media for growing sulfate reducers, methanogens, and various anaerobic chemolithoautotrophic and heterotrophic microorganisms that could use iron(III), manganese(IV), chlorate, or nitrate as an electron acceptor. To use larger sample volumes for inoculations and to avoid the dilution that a sediment sample undergoes during slurry preparation, core material freshly harvested with 5- and 60-mL syringes was also used directly for inoculations. All shipboard cultivation experiments and their media and incubation temperatures are shown in Table T7. In order to study the influence of sulfate as a limiting factor on the deep biosphere, both sulfate-free and sulfate-containing media were used for inoculation of samples from the sulfate-containing layers, the sulfate-methane transition zone, and sulfate-free layers. For details of the various media compositions and incubation conditions see "Procedures and Protocols" in "Microbiology" and Tables T4, T5, and T7, all in the "Explanatory Notes"
chapter.
13C substrate incubations were initiated for postcruise analysis by FISH-secondary ion mass spectrometry (SIMS) using material from Cores 201-1227A-3H and 5H and 201-1227D-4H. In this case, 10 mL of the master slurry was injected into each bottle. The 13C substrates used were methane, acetate, and glucose. For Cores 201-1227A-3H and 5H, one of each bottle was inoculated. For Core 201-1227D-4H, two bottles with each substrate and two 13C acetate bottles containing nonlabeled methane in the headspace were inoculated.
