SYNTHESIS OF PRIMARY FINDINGS
The sediments recovered during Leg 208 have shed new light on the nature of short-term paleoceanographic events of the last 70 m.y., while also improving our understanding of the general long-term trends as established by earlier cruises. Because of the exceptional core recovery in multiple holes and high-resolution core logging, we were able to resolve the complete spectrum of lithologic variability down to the centimeter scale, including orbitally paced oscillations. As a consequence, Leg 208, with the depth transect approach, was also able to establish the character of both the long- and short-term changes in sediment deposition in the vertical dimension, a constraint that is essential to understanding both the causes and consequences of paleoceanographic change. In the following section, we summarize the major findings of this leg to date, focusing primarily on those contributions that are novel or represent a significant improvement upon previous investigations.
Seismic profiles collected during the site survey cruise were used along with Leg 208 downhole logging and stratigraphic data to develop a three-dimensional reconstruction of sediment distribution on the northeastern flank of Walvis Ridge (Bartels et al., this volume). The multifrequency seismic data were collected using three seismic sources, two generator-injector (GI) guns (0.4 L; 100–500 Hz and 1.7 L; 30–200 Hz) and one water gun (0.16 L; 200–1600 Hz), in a quasi simultaneous mode in order to obtain the highest possible penetration and resolution of subsurface structures. Figure F4 shows a comparison between the GI gun data (spectrum 100–500 Hz) and the higher-resolution water gun data (spectrum 200–1600 Hz). Both data sets show the same seismic profile (GeoB01-035) crossing Site 1262. For comparison both profiles are plotted at the same scale. The images principally show the same sediment features in the vicinity of Site 1262 (i.e., parallel to subparallel layering of undisturbed pelagic sediments, a transparent, irregularly shaped body representing a debris flow or slump deposit within the uppermost 100 ms, and a sharp boundary at 6.45 s two-way traveltime representing the Eocene/Oligocene boundary). Closer examination, however, shows differences between these two data sets in terms of seismic facies. The water gun data show three distinct reflectors or reflector packages of sediments with higher amplitude beneath the sharp boundary at 6.45 ms. In contrast, the GI gun data suggest a gradual transition of physical properties. These sharp interfaces identified in the water gun data represent the Elmo horizon and the PETM and late Paleocene biotic event horizons (Röhl et al., 2004; Petrizzo, 2005), which are characterized by a density contrast of 10% in comparison to the Eocene and Paleocene sediments. This capability to image sediment layers on a meter to submeter scale was used to identify and correlate critical Cenozoic horizons in all of the Leg 208 cores.
Synthetic seismograms constructed using gamma ray attenuation (GRA) density data from downhole logs along with the shipboard stratigraphic data were used to identify, correlate, and date prominent seismic reflectors. The prominent reflectors all appear to correlate with major event horizons. Gridding of these reflectors or horizons was used to develop a paleoseafloor model that simulates sediment accumulation over the entire seismic grid for the last 65 m.y. (Bartels et al., this volume). This includes the response of sediment accumulation to changes in sediment fluxes in response to various processes such as scouring by bottom water currents. For example, the model shows evidence of a distinct change in bottom water current activity in the southeast sector of the study area between 28° and 30°S. Clearly visible is the development of a channel structure across Walvis Ridge, which is likely related to transport of North Atlantic Deep Water (NADW) and Antarctic Bottom Water between the Angola and Cape Basins (Fig. F5). Development of this channel indicates that the current strength increased toward the end of the middle Miocene, a phenomenon that is most likely related to key tectonic and climatic events during that period (e.g., Antarctic ice sheet expansion and closure of the Central American Seaway) (Nisancioglu et al., 2003).
The chronostratigraphy of the Leg 208 cores is based on bio-, magneto-, and cyclostratigraphic data. The biostratigraphic work carried out shipboard is reported in the Leg 208 Initial Reports volume (Zachos, Kroon, Blum, et al., 2004). Postcruise refinement of the biostratigraphy is still not completed and thus will not be discussed here.
Magnetostratigraphy
The soft, weakly magnetized carbonate sediments recovered during Leg 208 frequently produced erratic or seemingly biased directional records, at least in part resulting from coring-related deformation (Bowles, this volume). As a result, shipboard magnetostratigraphic interpretations were difficult or impossible to make over many intervals. Postcruise analysis of discrete samples allowed minor revision of several Paleocene to Upper Cretaceous reversal boundaries but unfortunately did little to refine most of the magnetostratigraphy.
Despite difficulty with the data, several polarity sequences were identifiable, including most of the major boundaries in the Pliocene–Pleistocene, an upper Miocene through Oligocene sequence at Sites 1265 and 1266, and an excellent Paleocene through Upper Cretaceous sequence at Sites 1262 and 1267. Although the inclination records from the Pliocene–Pleistocene are not very clean at most sites, we were frequently able to identify major reversal boundaries. However, assignment to a particular boundary was often aided in this interval by biostratigraphic datums or cyclostratigraphy. Of particular note, Chron C2n, close to the Pliocene/Pleistocene boundary, was identified at all sites except Site 1263. Sites 1265 and 1266 were combined to produce an interpretable upper Miocene through Oligocene sequence. In particular, this includes the excellent expression of Chron C6Cn at Site 1265 across the Oligocene/Miocene (O/M) boundary. This chron consists of three very distinctive short normal events which, combined with the biostratigraphic data and cyclostratigraphy, should allow refinement of the timescale across the O/M boundary. The Eocene was generally not well resolved at any of the sites, but an excellent Paleocene to Upper Cretaceous polarity sequence was recovered at Sites 1262 and 1267 (Fig. F6). At these two sites, the Paleocene cores were recovered either by APC or XCB systems in relatively indurated sediments with a higher clay content, which contributed to a lower degree of deformation over this interval. Identification of the upper and lower boundaries of Chron C24r is important for constraining the position of the Paleocene/Eocene (P/E) boundary within this chron. Whereas the lower boundary was identified shipboard, discrete sample analysis allowed placement of the upper boundary of Chron C24r at Sites 1262 and 1267; this constitutes a significant revision to the shipboard magnetostratigraphy at Site 1262. All remaining Paleocene and Upper Cretaceous reversal boundaries were identified at Sites 1262 and 1267, most to within ~10–30 cm.
Cycle Stratigraphy and Orbital Rhythms
Cyclic variations are present in core logging and core scanning data throughout the Maastrichtian–Holocene section at all sites. These variations are expressed by lithologic changes at a decimeter to meter scale. Cyclic variability was used during the cruise to correlate between parallel holes and to define a composite section for each site. At that time ~300 characteristic features (peaks or troughs) in magnetic susceptibility were identified and used to correlate between Leg 208 sites (Zachos, Kroon, Blum, et al., 2004). These tie points were dated using the age models of the individual sites. By adopting the average age of each tie point, a refined age model was constructed for the complete Leg 208 ~74-m.y. record. The distinct record of cyclic alternations in sediment physical properties provides high potential for refining the Neogene astronomical timescale and developing an astronomically tuned timescale of the Paleogene as far back as the Late Cretaceous (Westerhold et al., submitted [N1]). Detailed investigation of the sedimentary cycles and their relation to orbital forcing is an important objective of both already accomplished and ongoing postcruise studies. A definite tuning of the middle Eocene and older timescale faces fundamental issues because the precision of the orbital solutions (Laskar et al, 2004; Varadi et al., 2003) is limited, and there are also relatively large uncertainties in radiometric age constraints. However, a floating tuning and timescale may still be developed. Laskar et al. (2004) recommended that for construction of an astronomically calibrated timescale in the Paleogene only the very stable 405-k.y.-long eccentricity period should be utilized. Using filtering of various high-resolution core logs, including magnetic susceptibility and Fe intensity from X-ray diffraction (XRF), it was possible to identify primary cycles associated with eccentricity, both the 405- and 100-k.y. cycles. Within those cycles, the precession cycles could be easily identified and counted (Fig. F7) (e.g., Röhl et al., 2003, 2004; Westerhold et al., in press, submitted [N1]). Given the stability of the 405-k.y. eccentricity cycle, it was possible to obtain a best fit and thus derive numerical ages for the magnetochron boundaries and early Eocene warming events (Fig. F8) (Westerhold et al., in press, submitted [N1]; Röhl et al., 2006).
A major achievement of Leg 208 was the recovery of continuous undisturbed cores spanning several critical intervals or events. All events were recovered from at least two sites, and at least three of the early Cenozoic events were recovered at five sites. The composite sections along with the pronounced lithologic cycles allow each event to be observed in the context of orbitally paced oscillations in climate and ocean chemistry. The most prominent are the P/E and K/Pg boundaries, characterized by relatively rapid and extreme change. The other events, although less extreme, show characteristics that indicate brief extremes in climate and/or ocean carbonate chemistry. This includes the mid-Paleocene biotic event at 58.2 Ma (close to the lowermost occurrence of Heliolithus kleinpellii) (Bralower et al., 2002; Röhl et al., 2004; Petrizzo, 2005), the EOGM at 33.5 Ma, the early Oligocene Braarudosphaera layers at 28.5–30 Ma, and the early Miocene Bolivina acme of biserial foraminifers at ~18 Ma. In addition, several previously unrecognized events, characterized by clay layers similar to the P/E boundary but of a smaller scale, were identified in the upper Paleocene and lower Eocene of all sites (Zachos, Kroon, Blum, et al., 2004; Lourens et al., 2005; Röhl et al., 2005). The most distinct of these smaller events are two clay layers in the lower Eocene referred to as the Elmo and X events (Lourens et al., 2005; Röhl et al., 2005) events. The Elmo event occurs in uppermost Chron C24r, close to the uppermost occurrence of Discoaster multiradiatus. The assertion that these dissolution layers are linked to global events, rather than regional, is based on their presence in other ocean basins, primarily the Pacific, where they are documented in cores recovered from Shatsky Rise (Bralower, Premoli Silva, Malone, et al., 2002). The documentation of such events is of importance, as their occurrence was predicted on the basis of, for example, anomalous excursions in benthic foraminiferal assemblages (Thomas et al., 2000).
Cretaceous/Paleogene Boundary
The K/Pg boundary mass extinction reset the state of the global ecosystem. Marine K/Pg boundary sections thus provide an ideal opportunity for testing the recovery of global environment to biological disaster induced by a meteorite impact. Reduced surface ocean carbonate and organic matter production at the K/Pg boundary must have led to changes in chemical properties of the ocean and sedimentation patterns. The biogeochemical changes and associated sedimentation include (1) a drastic decrease in deep-sea carbonate sedimentation (Zachos and Arthur, 1986; D'Hondt and Keller, 1991), (2) a rapid decrease in export production, which should have led to decreased pelagic to benthic carbon isotopic gradients (Hsü et al., 1982; Stott and Kennett, 1989; Zachos et al., 1989; D'Hondt et al., 1998), and (3) migration of the lysocline or CCD. Note, however, that models evocating a collapse of primary productivity and/or the biotic pump are in disagreement with the lack of extinction of deep-sea benthic foraminifers (e.g., Culver, 2003; Thomas, in press).
Few completely recovered, well-dated marine K/Pg records are available to document the effects of biological disaster on sedimentation patterns. For example, although Zachos and Arthur (1986) showed that deep-sea carbonate accumulation did not recover for >2 m.y. after the mass extinction, a long-term, continuous record of deep-sea carbonate accumulation has only been generated for a single site (ODP Site 1001), where carbonate accumulation rates did not recover until 4 m.y. into the Paleogene (D'Hondt et al., 1998). Another example from drill holes (DSDP Leg 74) in the South Atlantic Ocean, Walvis Ridge, shows that carbonate accumulation and carbon isotope gradient recovery took millions of years (D'Hondt et al., 1996; Coxall et al., 2006). However, these records offer limited temporal resolution because of coring gaps and low-resolution chronologic control. With the benefit of the APC and employing a multiple-hole coring strategy, we were able to acquire a complete K/Pg section with the APC in two holes at Site 1262 (Walvis Ridge, Leg 208).
The boundary was cored at Sites 1262 and 1267. Double coring at these sites resulted in a total of four K/Pg records. The lithologic sequence in the K/Pg boundary interval is similar at both sites, as they differ in water depth by only 400 m. At both Sites 1262 and 1267, the K/Pg boundary records an abrupt transition from highly cyclic Maastrichtian clay-bearing nannofossil ooze with foraminifers (nannofossil Zone CC26) to overlying Paleocene dark reddish brown clay-rich foraminifer-bearing nannofossil ooze and nannofossil clay. This boundary coincides with a distinctive increase in magnetic susceptibility and a decrease in sediment lightness (Fig. F9), corresponding to an overall increase in the abundance of clays, oxides, and ash in the lowermost Paleocene. At Site 1262, microtectites are present at the boundary and overlying sediments grade upward into moderately bioturbated brown nannofossil- and foraminifer-bearing clay (foraminiferal Zones P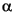 and P1a). Preliminary biostratigraphy shows the well-established abrupt change in plankton assemblages across the boundary at both sites (Luterbacher and Premoli Silva, 1964; Thierstein, 1982; Monechi, 1985). The white nannofossil ooze below the boundary yields diverse assemblages of the uppermost Maastrichtian Abathomphalus mayaroensis planktonic foraminiferal zone and nannofossil Zone CC26. The brown nannofossil- and foraminifer-bearing clay contains a high abundance of Woodringina hornerstownensis, Chiloguembelina midwayensis, and Chiloguembelina morsei as well as increasing abundance of Parvularugoglobigerina eugubina through the basal Paleocene (P). Based on this preliminary analysis, Zone P0 is not present at Walvis Ridge. However, this zone is absent at many deep-sea locations, and its absence may be ecologically determined rather than indicative of an unconformity (Norris et al., 1999). The substantial thicknesses of the uppermost Maastrichtian Micula prinsii Zone and the lowermost Danian P. eugubina Zone indicate that the K/Pg boundary is paleontologically complete. Moreover, the cycle stratigraphy is very robust with distinct spectral peaks in the precession and eccentricity bands. Thus, the Walvis Ridge sections provide a well-preserved and relatively detailed record of this major extinction event and the subsequent biotic recovery.
and P1a). Preliminary biostratigraphy shows the well-established abrupt change in plankton assemblages across the boundary at both sites (Luterbacher and Premoli Silva, 1964; Thierstein, 1982; Monechi, 1985). The white nannofossil ooze below the boundary yields diverse assemblages of the uppermost Maastrichtian Abathomphalus mayaroensis planktonic foraminiferal zone and nannofossil Zone CC26. The brown nannofossil- and foraminifer-bearing clay contains a high abundance of Woodringina hornerstownensis, Chiloguembelina midwayensis, and Chiloguembelina morsei as well as increasing abundance of Parvularugoglobigerina eugubina through the basal Paleocene (P). Based on this preliminary analysis, Zone P0 is not present at Walvis Ridge. However, this zone is absent at many deep-sea locations, and its absence may be ecologically determined rather than indicative of an unconformity (Norris et al., 1999). The substantial thicknesses of the uppermost Maastrichtian Micula prinsii Zone and the lowermost Danian P. eugubina Zone indicate that the K/Pg boundary is paleontologically complete. Moreover, the cycle stratigraphy is very robust with distinct spectral peaks in the precession and eccentricity bands. Thus, the Walvis Ridge sections provide a well-preserved and relatively detailed record of this major extinction event and the subsequent biotic recovery.
Preliminary results of postcruise work on the K/Pg boundary are shown in Figures F10 and F11 (Kroon et al., unpubl. data). High-resolution carbonate content and bulk carbon and oxygen isotope profiles show an abrupt change of dominant late Maastrichtian precessional cycles to marked early Paleocene short- and long-term eccentricity cycles (e.g., D'Hondt et al., 1996) at the K/Pg boundary (Fig. F10). The reason for the pronounced increase in the amplitude of the short- and long-term eccentricity frequencies at the K/Pg boundary is certainly related to the meteorite impact, although it is not yet clear which oceanic feedback processes responded to eccentricity forcing (D'Hondt et al., 1996). Carbonate accumulation shows that carbonate production at the K/Pg boundary ceased almost completely (Fig. F12). The bulk carbon isotope stratigraphy shows a large, abrupt shift toward negative values synchronous with the boundary and a more gradual change also toward negative values above the abrupt shift. The abrupt fall in values of ~1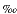 –1.5 at the boundary would represent eradication of the carbon isotope gradient in the water column in response to biological disaster in the pelagic realm (Kump, 1991; Zachos et al., 1989) in keeping with the large loss in carbonate production. The following gradual reduction in carbon isotope values is not observed in all carbon isotope records and thus difficult to reconcile.
–1.5 at the boundary would represent eradication of the carbon isotope gradient in the water column in response to biological disaster in the pelagic realm (Kump, 1991; Zachos et al., 1989) in keeping with the large loss in carbonate production. The following gradual reduction in carbon isotope values is not observed in all carbon isotope records and thus difficult to reconcile.
Bulk oxygen isotope stratigraphy shows the highest values of the entire late Maastrichtian–early Paleocene record just above the K/Pg boundary. This implies that surface waters of the South Atlantic cooled substantially, several degrees, for a period of several thousand years, although this phenomenon needs to be confirmed by oxygen isotope analysis of individual species of planktonic foraminifers.
The global CCD is thought to have deepened immediately following the extinction event (Zachos et al., 1989), in response to the sudden loss of carbonate production (Caldeira et al., 1990). Although various methods of documenting CCD change are available, we used the variability of the weight of the residue in the >63-µm fraction relative to the total weight (weight percent) as a proxy coupled with variability in carbonate content. The preservation state of the foraminifers in relation to this proxy was examined by using scanning electronic microscopy. Well-preserved foraminifers occurred synchronously with high weight percent of the residue in the >63-µm fraction. The profile of the weight percent of the residue in the >63-µm fraction shows that high weights are synchronous with relatively high percentage carbonate contents of the sedimentary record. Abrupt improvement of planktonic foraminiferal preservation was found uniquely at the K/Pg boundary, after which most of the variability oscillates at the frequencies of the short- and long-term eccentricity cycles.
Deep-sea benthic foraminifers at Site 1262 (as at other locations worldwide) did not undergo significant extinction. In contrast with some other locations, however (e.g., Alegret and Thomas, 2004, 2005), the Site 1262 record also shows no strong decrease in food delivery to the seafloor after the K/Pg boundary, with benthic foraminiferal accumulation rates indicative of a strongly fluctuating, at times very high, food supply (Alegret and Thomas, in press).
Mid-Paleocene Biotic Event
A prominent 10- to 30-cm-thick dark brown clay-rich calcareous nannofossil ooze was found at Sites 1262 and 1267, and a 10-cm-thick brown nannofossil chalk was found at Site 1266 (Röhl et al., 2004). These layers show a pronounced peak in magnetic susceptibility and XRF Fe data (Westerhold et al., submitted [N1]) that reflects an increase in clay content. Preliminary micropaleontological investigations suggest that this interval represents a short-lived event of considerable evolutionary significance. This interval corresponds to the P4 Globanomalina pseudomenardii planktonic foraminiferal zone and coincides with the evolutionary first occurrence of the nannofossil Heliolithes kleinpellii, an important component of upper Paleocene assemblages and a marker for the base of Zone CP5 (NP6) (lower upper Paleocene; ~58.2 Ma).
The event was also identified at Leg 113 Sites 689 and 690 (Thomas, 1990) and at Leg 198 Sites 1209–1212 (Bralower, Premoli Silva, Malone, et al., 2002; Petrizzo, 2005). Fundamental changes in faunal populations occur before, during, and after deposition of the clay-rich ooze. Planktonic foraminifers in the clay-rich layer are characterized by a largely dissolved low-diversity assemblage dominated by representatives of the genus Igorina (mainly Igorina tadjikistanensis) (Petrizzo, 2005). This low-diversity assemblage suggests some kind of oceanic perturbation of unknown origin. Together with the documented severe dissolution in this interval, the observed lithologic changes are likely to represent a response to increased seafloor carbonate dissolution caused by a transient shoaling of the lysocline and CCD. Regardless of origin, it is now clear from the high-resolution stratigraphy of the Leg 208 sites that this is a global event. Ongoing isotopic investigations should shed light on the nature of this extreme dissolution event (Röhl et al., 2004).
Paleocene/Eocene Thermal Maximum
The Paleocene/Eocene boundary was successfully recovered in multiple holes at five sites (e.g., Sites 1262, 1263, 1265, 1266, and 1267), spanning an estimated paleodepth range of ~1500 to ~3500 m. Shipboard physical property, lithologic, and biostratigraphic data indicate complete recovery of the Paleocene–Eocene transition interval at all sites except Site 1265 (see the Leg 208 Initial Reports volume; Zachos, Kroon, Blum, et al., 2004). Each boundary section is marked by a distinct clay layer. At the onset of the PETM, carbonate content plummets to ~0 wt%, producing a pronounced lithologic shift from nannofossil ooze to a clay interval that roughly doubles in thickness from ~20 to ~50 cm down the depth transect (Fig. F13). This general pattern of decreased carbonate content is present in all marine PETM records (e.g., Bralower et al., 1997; Thomas and Shackleton, 1996; Thomas, 1998; Thomas et al., 2000) and has been attributed to a massive methane flux to the ocean-atmosphere.carbon reservoir that elevated pCO2 levels, decreased carbonate ion concentrations, and shoaled the carbonate saturation profile (Dickens et al., 1995, 1997). Leg 208 records, however, clearly demonstrate that the South Atlantic paleo-CCD shoaled much more (>2000 m) than predicted by current models (~400 m) (Dickens et al., 1997), suggesting release of a much larger volume of carbon.
Biostratigraphically, the onset of clay deposition coincides with the highest occurrence of the benthic foraminifer Stensioeina beccariiformis and other typical upper Paleocene taxa (Fig. F13). Calcareous microfossils are absent to extremely rare and generally poorly preserved in the lowermost clay, reflecting some combination of deleterious benthic conditions, decreased carbonate export production, and intensified carbonate dissolution. Lowermost Eocene benthic foraminifers near the base of the clay layer (Fig. F13) are extremely low in abundance and diversity and minute in size. Aragonia aragonensis, Tappanina selmensis, Oridorsalis umbonatus, and Bulimina spp. dominate at shallower Sites 1263, 1265, and 1266, whereas abyssaminids and clinapertinids are common at these sites but dominate at deeper Sites 1262 and 1267 and in the lowermost sample at the shallower sites. In general, assemblage composition in the clayey intervals is strongly variable. Nannofossils are common and show only slight dissolution. The last appearance of the planktonic foraminifer Morozovella velascoensis roughly coincides with the onset of the PETM, and no related "excursion" taxa (e.g., Morozovella allisonensis, Acarinina sibaiyaensis, and Acarinina africana) occur within the PETM—a biogeographic pattern that stands in stark contrast to those documented at lower and higher paleolatitudes (Kelly et al., 1998; Kelly, 2002). Planktonic foraminifers within the clay layer primarily consist of extremely rare and poorly preserved specimens of Acarinina soldadoensis. Nannofossil assemblages within the PETM clay are markedly poorer in preservation, lower in abundance and richness, and dominated by discoasters.
The base of the PETM recovery interval may be defined as the onset of increasing carbonate content, which produced a gradational upsection lithologic sequence at each site of nannofossil-bearing clay, nannofossil clay, clay-bearing nannofossil ooze, and, finally, nannofossil ooze. Recovery intervals are thicker at shallower sites, likely reflecting higher overall mass accumulation rates coupled with the time-transgressive deepening of the carbonate saturation profile. Magnetic susceptibility values are consistent with this scenario, with shallower sites showing more oscillations within the generally decreasing trends. As carbonate content increased through the PETM recovery interval, planktonic foraminiferal preservation improved and faunal abundance and richness increased to include morozovellids, acarininids, subbotinids, and rare globanomalinids. The first occurrences of the nannofossils Rhomboaster cuspis and Rhomboaster calcitrapa, basal members of the Rhomboaster-Tribrachiatus lineage, are within the recovery interval and provide potentially isochronous biomarkers for intersite correlation. Above these nannofossil first occurrences, three prominent bioevents occur in varying stratigraphic order at all five sites: (1) benthic foraminiferal compositions shift from lowermost Eocene low-diversity diminutive assemblages to lower Eocene moderate-diversity assemblages, (2) nannofossil assemblages show a marked relative decrease in Fasciculithus spp., and (3) nannofossil assemblages also show a marked relative increase in Zygrhablithus bijugatus. This relative increase in Z. bijugatus typically coincides with the benthic foraminiferal extinction event (BEE) in other regions (e.g., Shatsky Rise and central Pacific) but occurs later at Walvis Ridge and may be correlative with the Fasciculithus–Rhomboaster abundance reversal reported at equatorial Pacific Sites 1220 and 1221 (Lyle, Wilson, Janecek, et al., 2002). These intersite differences in bioevent ordination may represent incomplete sampling coverage or real paleoenvironmental diachroneity between sites.
A number of postcruise investigations were undertaken to identify the impacts of the PETM on regional climate, ocean chemistry and ecology, and sediment accumulation. These investigations include (1) establishing high-resolution isotope and geochemical stratigraphies for improving site to site correlations and computing carbonate accumulation rates (Zachos et al., 2005; Röhl et al., submitted [N2]), (2) developing an organic compound–specific carbon isotope record (Hasegawa et al., this volume), (3) establishing changes in the general mineralogy and grain size distribution of the nonsoluble sediment fraction (Nicolo and Dickens, this volume), and (4) evaluating the local ecological impacts on phytoplankton and benthic foraminifers.
High-Resolution Stable Isotope Stratigraphies
Bulk carbon isotope records have proved to be useful for first-order correlation of marine P/E boundary sections. Each of the boundary sections in the Leg 208 sites was sampled at 1- to 5-cm intervals for the purpose of developing high-resolution isotope stratigraphies across the clay layers. These samples were supplemented with continuous channel samples (quarter and half-rounds) collected postcruise. The stable isotope analyses were carried out in four different laboratories (University of California, Santa Cruz, University of Florida, Bremen University, and Frjei University) (Zachos et al., 2005).
Each of the five bulk isotope records show nearly identical patterns over a 3- to 4-m interval spanning the boundary, with a prominent negative carbonate isotope excursion (CIE) coeval with the clay layers (Fig. F14). The thickness of the excursion layers and magnitudes of the CIE, however, systematically decrease with depth, implying a preservation bias. Site 1263 shows the largest CIE of about –3.0, whereas Site 1262 has a –2.0 excursion. This pattern is attributed to the combined effects of dissolution and mixing of preexcursion carbonate in the clay layers. Excluding these low-carbonate intervals, the carbon isotope stratigraphies were used to correlate the five sites to each other as well as to ODP Site 690 (Zachos et al., 2005). This correlation provided the primary basis for determining the duration of deposition of the clay layer, ~40 k.y., and the duration of the time lag for carbonate deposition to resume at the deepest site relative to the shallowest, and hence, the recovery time for the CCD.
The timescale for recovery of the CCD recorded in the Leg 208 cores is consistent with general carbon cycle theory. In simulations of the ocean carbon system response to a rapid (~300 yr) anthropogenic CO2 pulse, a release of 4500 Gt C results in severe carbonate undersaturation over most of the deep sea for a period exceeding 10 k.y. (Archer et al., 1997). Saturation levels did not recover for 40 k.y., and it took more than 100 k.y. for most of the excess carbon to be sequestered. Moreover, the restabilization of carbonate content and magnetic susceptibility values slightly higher and lower, respectively, than their pre-PETM values, are also consistent with theory. This pattern, which is present at other PETM sites (e.g., Southern Ocean Site 690), occurs in model simulations as a transient lysocline overdeepening in response to oceanic mass balancing of increased bicarbonate, carbonate, and Ca2+ concentrations (by way of enhanced chemical weathering and runoff) (Dickens et al., 1997).
Compound-Specific Carbon Isotope Stratigraphies
Another precruise objective was to determine whether the Paleocene and Eocene sediments on Walvis Ridge contained sufficient quantities of marine organic matter to isolate specific organic compounds, such as alkenones, for stable carbon isotope analysis. Such a record could potentially be used to reconstruct changes in pCO2 through the PETM. Shipboard gas chromatographic (GC) analyses of 5-cm quarter-round samples collected from sediments spanning the clay layer revealed the near-complete absence of alkenones and instead the presence of terrestrially derived n-alkanes. Based on these results, a decision was made to generate a detailed compound-specific carbon isotope record across the P/E boundary. Quarter-round samples, 5 cm in length, were collected from the shallowest Site 1263 during the postcruise sampling party. For GC-mass spectrometry (MS) analysis, the samples were prepared following standard techniques to isolate insoluble organic residues (Hasegawa et al., this volume). Three heavy n-alkanes, nonacosane (n-C29), hentriacontane (n-C31), and tritriacontane (n-C33), were isolated in sufficient quantities for carbon isotope analysis. The  13C values range from –26 to –33 (Fig. F15). The record shows a prominent ~5 negative excursion coincident with the base of the clay layer. The isotope profiles of the three normal alkanes are similar to that of carbonate carbon, though the magnitude of the excursion is larger, roughly 4–5. In addition, the n-alkane records show an inflection at 334.70 meters composite depth (mcd), which is roughly equivalent to the CIE in the bulk carbonate record.
13C values range from –26 to –33 (Fig. F15). The record shows a prominent ~5 negative excursion coincident with the base of the clay layer. The isotope profiles of the three normal alkanes are similar to that of carbonate carbon, though the magnitude of the excursion is larger, roughly 4–5. In addition, the n-alkane records show an inflection at 334.70 meters composite depth (mcd), which is roughly equivalent to the CIE in the bulk carbonate record.
These long-chain n-alkanes are a common component of terrestrial vegetation. The low carbon isotope values are consistent with this interpretation, indicating they were derived from C-3 plants, most likely in South Africa. The heavy n-alkanes are a common component of leaf waxes, which tend to be highly resistant to degradation, a possible reason for the high abundances in the Leg 208 sediments relative to compounds from marine organic matter. The magnitude of the excursion is considerably larger than that recorded in carbonates from these same cores. This particular finding is significant, as it is consistent with observations of other terrestrial carbon isotope records which typically show large excursions, on the order of 5–6, or nearly twice that of marine records (Koch et al., 1992; Bowen et al., 2004; Pagani et al., 2006). Several models have been proposed to explain the differing magnitudes of the marine and terrestrial CIEs including (1) truncation of the marine carbonate records by dissolution and bioturbation (Zachos et al., 2005), (2) damping of the marine signal by lower pH and CO3 content, and (3) amplification of terrestrial vegetation signal because of higher pCO2 and/or humidity (i.e., Bowen et al., 2004). It is also possible that the source of n-alkanes (i.e., type of plants) changed during the PETM. Because this is the first time that both the marine and terrestrial signals were reconstructed from the same core, we can eliminate bioturbation as the primary source of the reduced amplitude of the marine records.
Another important feature of this record is the relative 13C offsets between the three groups of analyzed n-alkanes. In general, the heavier n-alkanes (n-C33) tend to have consistently lower 13C values than the lighter, similar to the pattern in modern plants. This intercompound fractionation offset, however, decreases during the PETM, implying some change in plant metabolism/physiology. Whether this change in fractionation was caused by the effects of higher pCO2, humidity, species composition, or metabolic rates is unknown.
Calcareous Algae
Large shifts in the relative abundances of calcareous algae have been recorded in all marine P/E boundary sections studied to date (e.g., Monechi et al., 2000; Bralower, 2002; Tremolada and Bralower, 2004; Raffi et al., 2005; Gibbs et al., 2006). Similarly, large shifts occur in the Leg 208 sites. Some of the changes are clearly related to dissolution, whereas other shifts appear to reflect actual paleoecological responses. Given the recent interest in the potential impacts of ocean acidification on marine calcifiers, particularly phytoplankton, several postcruise studies were undertaken to ascertain changes in nannofossil assemblages through this event.
Initial results reveal several important patterns involving several major groups of nannofossils including genera Fasciculithus, Rhomboaster, Tribrachiatus, and Discoaster (Monechi and Angori, this volume; Agnini et al., submitted [N3]). At the deeper Sites 1262 and 1266, it is clear that dissolution strongly influenced abundance patterns. In particular, the abundances of dissolution-resistant taxa such as Rhomboaster are much higher than at the shallowest site. As such, we will only discuss patterns observed at the shallowest Site 1263 where preservation is moderate to good throughout. Only in a 6-cm interval (between 335.68 and 335.62 mcd), corresponding to an earlier stage of the CIE onset, do assemblages show the effect of dissolution that, nevertheless, does not significantly alter the assemblages in terms of taxonomic composition. The recorded biostratigraphic signals are diverse and involve different components of the nannofossil assemblages. The changes can be divided into successive biohorizons, marked by increased/decreased abundances of taxonomic groups, sudden appearances, or ranges of individual taxa, all of which are described below and labeled as Biohorizons N1–N5 in Figure F16.
The N1 biohorizon is coincident with the onset of the CIE and is defined by a drastic decrease in diversity and abundance of the genus Fasciculithus. The turnover involved the characteristic large species in fasciculith assemblages of the uppermost Paleocene, like Fasciculithus richardii, F. schaubii, F. hayi, F. toni, and F. mitreus, that suddenly disappeared. This event is recorded in Paleocene/Eocene boundary sections across the globe (Backman, 1986; Monechi et al., 2000; Raffi et al., 2005). At Site 1263, the N2 biohorizon coincides with the second step in the onset of the CIE, where survivor species of fasciculiths, Fasciculithus alanii, F. tympaniformis, and F. involutus, peak in abundance, with the interval of peak abundance in Fasciculithus correlated to the lowest diversity of benthic foraminifers. Also, the first occurrence of F. thomasii is recorded, closely followed by the appearance of the peculiar forms of asymmetrical and deformed discoasterids. These discoasterids show an asymmetrical radial structure with a prominent irregular central knob. The first occurrence of rare, small specimens of Discoaster salisburgensis further characterizes this interval. The occurrences of the two short-lived and deformed discoasterids further delimit the N2 biohorizon. The N3 biohorizon is marked by the base of the Rhomboaster calcitrapa group. Within the CIE, Rhomboaster morphotypes belonging to the spined R. calcitrapa group and R. cuspis have the lowermost occurrence. R. calcitrapa group specimens have been observed in most of the known P/E sections and appear to represent a globally distributed evolutionary event. In the upper part of the CIE, at the beginning of the "recovery interval" where bulk 13C values start to increase and weight percent CaCO3 content further increases (exceeding 60 wt% at Site 1263), we record a sharp decrease of genus Fasciculithus and an increase in the genus Zygrhablithus. This crossover in abundance Fasciculithus vs. Zygrhablithus defines biohorizon N4. Because this event is consistently recorded close to the P/E boundary in different oceanic settings (Bralower, 2002; Tremolada and Bralower, 2004; Gibbs et al., 2006), it is clearly related to PETM environmental changes. Zygrhablithus is likely occupying the ecological niche vacated by the permanent global decline of Fasciculithus (Bralower, 2002). The interval between Biohorizons N4 and N5 coincides with the recovery phase in bulk 13C, and it is also characterized by increases in the relative abundances of Discoaster and Sphenolithus. Biohorizon N5 is delineated by the uppermost occurrence of the spined Rhomboaster spp. An increase in abundance of Tribrachiatus bramlettei is recorded at the last occurrence of spined Rhomboaster. Biohorizon N5 is correlative with the full recovery of carbonate content at all sites, when CaCO3 percentage stabilized at ~90 wt% (Zachos et al., 2005) (Figs. F14, F16). The high abundances of this particular group may be a consequence of the highly oversaturated state of the ocean.
Terrigeneous Sediment Flux
The terrigenous fraction of lower lower Paleogene sediments is likely composed of both hemipelagic and eolian material at all Leg 208 sites. Terrigenous component-specific grain-size distributions (Fig. F17) indicate that Site 1267, positioned on a deeper flank of Walvis Ridge, received a greater flux of hemipelagic sediment than did the relatively shallow near-crest Site 1263 during this interval (Nicolo and Dickens, this volume). This observation is only possible because of the unique geologic setting of Walvis Ridge and the depth transect strategy of Leg 208 and is consistent with both previous work at DSDP Site 527 (Rea and Hovan, 1995) and general views of open-ocean terrigenous deposition (e.g., Rea, 1994). These results imply that an early Paleogene eolian grain-size record may be gleaned from Site 1263 samples (Nicolo and Dickens, this volume).
Elmo (Eocene Thermal Maximum-2)
At ~20–35 m above the P/E boundary, a red-colored 5- to 15-cm carbonate-depleted layer was found at all sites with lower Eocene strata (Fig. F18). During the cruise this layer was tentatively placed in Chron C24n and called the Chron C24n event. Subsequent postcruise work suggests the layer may lie just below Chron C24n (Bowles, this volume). Moreover, a second name, Elmo, was applied to the event. The Elmo shows similar color characteristics to the P/E boundary layer as well as a drop in calcium carbonate content and an increase in magnetic susceptibility and natural gamma radiation (NGR) values, but these changes are not as significant. The Elmo is characterized by a double peak in the 1-cm sampled point magnetic susceptibility records of all sites, which are marked by events a and b in Figure F18. At the shallowest sites (Sites 1263–1266), events a and b are most distinctly developed, whereas at the deepest sites (Sites 1267 and 1262) they are merged. At the latter sites, an additional peak in magnetic susceptibility values is observed slightly above the main clay layer. At middepth Site 1266, a thin white-colored layer is found immediately above the red-colored horizon. The uppermost occurrence of nannofossil D. multiradiatus is recorded close to the Elmo. This layer, which appears to be present in the magnetic susceptibility records of sites drilled on Shatsky Rise, is associated with benthic foraminiferal assemblages similar in character to those of the PETM. This implies a transient shift of paleoenvironmental conditions toward those documented for the PETM.
Postcruise studies of the Elmo include stable isotope analyses of bulk sediment and foraminifers collected from samples taken at 1-cm spacing across the event. This work reveals negative carbon and oxygen isotope excursions in both bulk and planktonic and benthic foraminifer samples coeval with the low-carbonate layer (Fig. F19) (Lourens et al., 2005). The magnitude of the carbon isotope excursion, 1.5–2.0, is roughly half that of the PETM. In addition, the onset seems more gradual. The oxygen isotope excursion in benthic foraminifers is on the order of 0.8, suggesting 2°–3°C warming, again about 30%–40% of that observed for the PETM. As discussed above, initial cycle stratigraphic analysis of the magnetic susceptibility data suggest that this event lies within five 400-k.y. eccentricity cycles of the PETM, or about 2 m.y. younger (Lourens et al., 2005), though subsequent work involving core scanned XRF data suggests a slightly shorter duration (Westerhold et al., in press). Given the similarity of key attributes of these hyperthermals (e.g., transient warming, benthic foraminiferal assemblage changes, and CIE), we designate the PETM as Eocene Thermal Maximum 1 (ETM-1), and the Elmo as Eocene Thermal Maximum 2 (ETM-2).
X Event (Eocene Thermal Maximum-3)
We document a third thermal maximum in lower Eocene (~52 Ma) sediments. The prominent clay layer, named the "X" event, occurs in planktonic foraminifer Zone P7 and calcareous nannofossil Zone CP10 at four sites. Benthic foraminifers have low diversity and high dominance and are dominated by small individuals of Nuttallides truempyi and various abyssaminids, resembling the post-PETM extinction assemblages, with more severe effects at deeper sites. Calcareous nannofossil assemblages show similar trends to the PETM, with major changes in the genera Discoaster and Zygrhablithus, but with differences in magnitudes and fluctuations. High-resolution bulk carbonate stable isotope values at relatively shallow Site 1265 show a rapid, 0.6 drop in 13C and 18O, followed by an exponential recovery to preexcursion values (Röhl et al., 2005; Röhl et al., submitted [N2]), a pattern similar to that of the PETM and Elmo. Planktonic foraminiferal 13C values (Morozovella subbotina and Acaranina soldadoensis) in the deepest Site 1262 decrease by 0.8–0.9, those of the benthic foraminifer Nuttalides truempyi by 1. Evaluations for mechanisms for the widespread change in deepwater chemistry, its connection to the surface water response, and the temporal relation of the event, as well as the PETM and Elmo, with current astronomical solutions are still ongoing.
Eocene/Oligocene Boundary and Early Oligocene Glacial Maximum
Sediment recording the response of South Atlantic Ocean to global cooling and CCD deepening during the Eocene–Oligocene transition was recovered across a broad range of depths on the northeastern flank of Walvis Ridge during Leg 208, but the transition is not well characterized biostratigraphically. The shallower Sites 1263 and 1265 contain the most complete records, but even at these sites, calcareous nannofossil and benthic and planktonic foraminiferal assemblages are affected by dissolution, downslope transport, and reworking.
Calcareous nannofossil preservation in the Eocene/Oligocene (E/O) boundary interval is moderate because of dissolution, etching, and common reworking. Nannofossil events that bracket the boundary are the uppermost occurrence of the rosette-shaped discoasters (last occurrence [LO] of Discoaster barbadiensis at 34.2 Ma and LO of Discoaster saipanensis at 34.0 Ma) and that of Ericsonia formosa (32.9 Ma). The LO of D. saipanensis and the LO of E. formosa could be observed at all sites, but these are uppermost occurrences and thus are prone to reworking. The bottom of the increase in the abundance of Ericsonia obruta is at the E/O boundary (33.7 Ma) but was observed at Site 1263 only. Fortunately, the global increase in 18O values in deep-sea benthic foraminifers (Oi-1; ~33.6 Ma), occurring within the interval between the LO of D. saipanensis and the LO of E. formosa (34.0–32.9 Ma) (e.g., Zachos et al., 2001), is recovered clearly in the shallow Sites 1263, 1265, and 1266 (Fig. F20). At the deepest Site 1262, carbonate fossils were completely dissolved below the E/O boundary at 33.7 Ma (Fig. F20) (Liu et al., 2004b), and the lithologic change from brown clay below to light brown to gray nannofossil ooze or foraminifer-bearing nannofossil ooze above occurs abruptly over a ~0.5-m interval.
At all sites, a distinct increase in carbonate occurs starting from the E/O boundary (Fig. F21). This increase is associated with the strongest foraminifer dissolution, leading the EOGM by ~100 k.y. The increased carbonate content across the boundary interval indicates that the lysocline and CCD deepened substantially and rapidly at the Walvis Ridge transect sites. In the latest Eocene, the lysocline was between the paleodepths of Sites 1266 and 1267. During the Eocene–Oligocene transition, the lysocline/CCD deepened abruptly to a depth below that of Site 1262, at least 600 m deepening. The recovery of foraminiferal preservation on the basis of coarse fraction (carbonate grain size > 63 µm) lags by ~400 k.y. (Fig. F21), implying that the shift in carbonate preservation was triggered by changes in climate. This significant downward shift in lysocline/CCD in the E/O boundary interval has also been observed in other ocean basins (e.g., Zachos et al., 1996; Lyle, Wilson, Janecek, et al., 2002) and possibly reflects an increase in mechanical and chemical weathering rates on continents and related changes in ocean chemistry associated with global cooling (EOGM). The peak in carbonate content, however, is transient, as values decline shortly thereafter. This peak in carbonate value corresponds with a magnetic "normal" that may represent Chron C13n.
Early Miocene High Abundance of Bolivinid Biserial Foraminifers
An enigmatic feature of early Miocene oceanography was the so-called "high abundance of bolivinids (HAB) event" (Thomas, 1986, 1987; Smart, 1992; Smart and Murray, 1994; Smart and Ramsay, 1995). During this event (~18.9–17.2 Ma) biserial foraminifers (assigned to the benthic genus Bolivina) were abundant at bathyal to abyssal depths in the eastern Atlantic and western Indian Ocean but were not observed in the western equatorial Atlantic Ocean, eastern equatorial Pacific Ocean (Thomas, 1985), or the Weddell Sea (Thomas, 1990), although the sediments in the correct size fraction and of the correct age were studied. The HAB event was enigmatic: Thomas (1986, 1987), Smart (1992), and Smart and Murray (1994) speculated that it might represent a period of low oxygen conditions during sluggish circulation. Smart and Ramsay (1995) argued that the event was associated with an oxygen-depleted water mass restricted to the eastern Atlantic and western Indian Oceans. In the modern oceans, however, such high relative abundances of bolivinids occur only where a severe oxygen minimum zone impinges on the seafloor (e.g., Bernhard and Sen Gupta, 1999), and there is no evidence for dysoxia (e.g., lamination or high organic carbon content) during the HAB event.
Smart and Thomas (2006) used morphological and stable isotope data of biserial foraminifers in samples (21.0–15.8 Ma) from Sites 1264 and 1265 to document that the early Miocene biserial forms are not benthic but planktonic and should be assigned to the genus Streptochilus. These biserial forms may have bloomed opportunistically, as suggested by their small size and high abundance, as also inferred for Paleogene biserial planktonic foraminifers (e.g., Hallock et al., 1991). During the interval of their high abundance, however, the high primary productivity did not result in high export productivity to the seafloor. Benthic foraminiferal accumulation rates have been described as a proxy for productivity (Herguera and Berger, 1991) but are low in the interval with highest abundance of biserials at Walvis Ridge and Site 608 (Diester-Haass and Billups, 2005). The low export productivity might be explained by the presence of a deep thermocline, in which regeneration rates of organic matter are high (e.g., Fischer et al., 2003), existing over a large geographic area.
The question remains: Why was there widespread, active, but intermittent upwelling of nutrient-rich waters in eastern Atlantic and western Indian only at that time (18.9–17.2 Ma)? The cause might be sought in circulation changes: Northern Component Water (NCW) may have been present between 19 and 17 Ma, its formation possibly triggered by the sinking of the Greenland-Scotland Ridge (Wright, 1998). Another possible trigger for NCW formation might be the development of a deep ACC at ~19 Ma (e.g., Anderson and Delaney, 2005), leading to the formation of NCW (Sijp and England, 2004). A deep ACC is necessary for strong vertical mixing in the southern oceans, which leads to increased nutrient contents in upwelled waters such as in the Subantarctic Mode Waters, which presently support global productivity (Sarmiento et al., 2004). The gradual closing of the Mediterranean to the Indian Ocean (Harzhauser et al., 2002) may have increased the flow of surface waters from the Indian Ocean into the Atlantic around South Africa (Agulhas Leakage), leading to higher density waters in the North Atlantic (Weijer et al., 1999) and local deep convection (de Ruijter et al., 2006). Active deep convection in a deep ACC and in the Agulhas Leakage in the southern oceans thus could have provided the nutrient-rich waters upwelled in the eastern Atlantic and western Indian Ocean.
But if that were the case, then why did the high abundance of biserial planktonic foraminifers end at ~17.5 Ma? Possibly, vigorous North Atlantic deep circulation ended somewhat earlier than envisaged by Wright (1998), thus ending vigorous upwelling in the eastern Atlantic Ocean. It is, however, not clear why the high abundances of biserial planktonic foraminifers did not resume in the middle Miocene, with increased cooling and formation of NCW. Possibly, upwelling at that later time became more constrained to its modern locations rather than more widespread because of the overall increase of circulation vigor during the middle Miocene cooling.
Cenozoic Seawater Chemical Evolution
Several postcruise studies used fossils obtained from the Leg 208 cores to establish variations in seawater chemistry, specifically radiogenic Nd, Sr, and Li isotope distributions during key intervals of the Cenozoic. In the case of Nd, the objective was to establish changes in deep ocean circulation patterns, whereas the Sr investigation was oriented toward establishing changes in geochemical fluxes. Li isotopes were used along with Li/Ca data to evaluate changes in the character of chemical weathering of continental rocks.
Modern thermohaline circulation, with vigorous deepwater formation in the North Atlantic, was established some time in the middle Cenozoic (Wright and Miller, 1993). Traditionally, carbon isotope distributions as recorded in benthic foraminifers between the deep Pacific and Atlantic were used to established general circulation patterns. However, this approach has limited sensitivity during times when nutrient inventories were lower and/or productivity was suppressed. To establish the timing of NADW formation, Via and Thomas (2006) reconstructed changes in bottom water Nd isotope ratios in the South Atlantic during the Oligocene using samples from the Leg 208 depth transect. Bottom water Nd isotope ratios were obtained from analysis of fish teeth, which acquire Nd concentrations after deposition on the seafloor. Using the Nd isotope data from Walvis Ridge along with data from sites in the North Atlantic and Southern Ocean, they found an increase in the north-south Nd isotope gradient, suggesting the initial transition to a bipolar mode of deepwater circulation occurred in the early Oligocene, at ~33 Ma (Fig. F22). They attribute the onset and strengthening of deepwater production in the North Atlantic to tectonic deepening of the sill separating the Greenland-Norwegian Sea from the North Atlantic (Wright and Miller, 1996).
The late Paleocene–early Eocene warming trend has been attributed to increased volcanism and CO2 outgassing, most likely associated with North Atlantic rifting. In theory, such volcanism should have lowered the Sr isotopic composition of seawater. To test this, Hodell et al. (submitted [N4]) constructed a high-resolution Sr isotope record over the upper Paleocene and lower Eocene using foraminifers from Leg 208 cores (Fig. F23). The sampling interval, ~50 k.y., is the highest such for this time interval and therefore reveals features not obvious in lower-resolution records. The main feature is a subtle, steady decline in 87Sr/86Sr over the late Paleocene and early Eocene, followed by a slight rise through the middle Eocene. The decline coincides with a long-term gradual warming that culminates in the EECO. This pattern of decreasing 87Sr/86Sr coeval with increased warming has been documented in the Cretaceous (e.g., Jones and Jenkyns, 2001) and has been attributed to increased flood basalt volcanism and CO2 outgassing. Thus, the Leg 208 87Sr/86Sr record implies a similar connection between volcanism and climate in the Paleogene.
The concentration of Li in the ocean is thought to be controlled by variations in hydrothermal fluxes. Using planktonic foraminifers from Site 1264 along with samples from several other ODP cores, Hathorne and James (2006) reconstructed Li variations for the Atlantic and Pacific ocean over the last 18 m.y. The 7Li records for the Atlantic and Pacific look identical, suggesting that the patterns reflect changes in mean ocean Li (Fig. F24). Their findings also suggest very little change in Li/Ca ratios. By assuming that the hydrothermal flux of Li into the oceans and the flux of Li removed from the oceans during low-temperature uptake by marine basalts and sediments have not changed significantly since 18 Ma and using published records for the seawater Ca concentration, the seawater Li/Ca and 7Li records are used to estimate global average river 7Li and Li fluxes. This analysis requires a decrease in the riverine flux of dissolved Li between 16 and 8 Ma but an increase in 7Li value of the riverine input. These data imply that both silicate weathering rates and weathering intensity decreased over this interval, which may have contributed to changing levels of atmospheric CO2. In contrast, the computations indicate riverine flux of Li has increased since 8 Ma while 7Li has increased. This implies that the silicate weathering rate has increased, while weathering intensity has decreased.
